 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4v5m | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | tRNA tranlocation on the 70S ribosome: the pre-translocational translocation intermediate TI(PRE) | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | RIBOSOME / TRANSLATION / ELONGATION CYCLE | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報ribosome disassembly / translation elongation factor activity / large ribosomal subunit / ribosome binding / regulation of translation / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / transferase activity ...ribosome disassembly / translation elongation factor activity / large ribosomal subunit / ribosome binding / regulation of translation / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / transferase activity / 5S rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / tRNA binding / cytoplasmic translation / rRNA binding / negative regulation of translation / ribosome / structural constituent of ribosome / ribonucleoprotein complex / translation / GTPase activity / mRNA binding / GTP binding / protein homodimerization activity / zinc ion binding / metal ion binding / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||||||||
| 生物種 |   THERMUS THERMOPHILUS (バクテリア) THERMUS THERMOPHILUS (バクテリア) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 7.8 Å | ||||||||||||
 データ登録者 データ登録者 | Ratje, A.H. / Loerke, J. / Mikolajka, A. / Bruenner, M. / Hildebrand, P.W. / Starosta, A.L. / Doenhoefer, A. / Connell, S.R. / Fucini, P. / Mielke, T. ...Ratje, A.H. / Loerke, J. / Mikolajka, A. / Bruenner, M. / Hildebrand, P.W. / Starosta, A.L. / Doenhoefer, A. / Connell, S.R. / Fucini, P. / Mielke, T. / Whitford, P.C. / Onuchic, J.N. / Yu, Y. / Sanbonmatsu, K.Y. / Hartmann, R.K. / Penczek, P.A. / Wilson, D.N. / Spahn, C.M.T. | ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2010 タイトル: Head swivel on the ribosome facilitates translocation by means of intra-subunit tRNA hybrid sites. 著者: Andreas H Ratje / Justus Loerke / Aleksandra Mikolajka / Matthias Brünner / Peter W Hildebrand / Agata L Starosta / Alexandra Dönhöfer / Sean R Connell / Paola Fucini / Thorsten Mielke / ...著者: Andreas H Ratje / Justus Loerke / Aleksandra Mikolajka / Matthias Brünner / Peter W Hildebrand / Agata L Starosta / Alexandra Dönhöfer / Sean R Connell / Paola Fucini / Thorsten Mielke / Paul C Whitford / José N Onuchic / Yanan Yu / Karissa Y Sanbonmatsu / Roland K Hartmann / Pawel A Penczek / Daniel N Wilson / Christian M T Spahn /  要旨: The elongation cycle of protein synthesis involves the delivery of aminoacyl-transfer RNAs to the aminoacyl-tRNA-binding site (A site) of the ribosome, followed by peptide-bond formation and ...The elongation cycle of protein synthesis involves the delivery of aminoacyl-transfer RNAs to the aminoacyl-tRNA-binding site (A site) of the ribosome, followed by peptide-bond formation and translocation of the tRNAs through the ribosome to reopen the A site. The translocation reaction is catalysed by elongation factor G (EF-G) in a GTP-dependent manner. Despite the availability of structures of various EF-G-ribosome complexes, the precise mechanism by which tRNAs move through the ribosome still remains unclear. Here we use multiparticle cryoelectron microscopy analysis to resolve two previously unseen subpopulations within Thermus thermophilus EF-G-ribosome complexes at subnanometre resolution, one of them with a partly translocated tRNA. Comparison of these substates reveals that translocation of tRNA on the 30S subunit parallels the swivelling of the 30S head and is coupled to unratcheting of the 30S body. Because the tRNA maintains contact with the peptidyl-tRNA-binding site (P site) on the 30S head and simultaneously establishes interaction with the exit site (E site) on the 30S platform, a novel intra-subunit 'pe/E' hybrid state is formed. This state is stabilized by domain IV of EF-G, which interacts with the swivelled 30S-head conformation. These findings provide direct structural and mechanistic insight into the 'missing link' in terms of tRNA intermediates involved in the universally conserved translocation process. | ||||||||||||
| 履歴 |
| ||||||||||||
| Remark 700 | SHEET DETERMINATION METHOD: DSSP THE SHEETS PRESENTED AS "QA" IN EACH CHAIN ON SHEET RECORDS BELOW ... SHEET DETERMINATION METHOD: DSSP THE SHEETS PRESENTED AS "QA" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 6-STRANDED BARREL THIS IS REPRESENTED BY A 7-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. |
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4v5m.cif.gz | 3.2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4v5m.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 4v5m.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 4v5m_validation.pdf.gz | 1.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 4v5m_full_validation.pdf.gz | 4.5 MB | 表示 | |
| XML形式データ | 4v5m_validation.xml.gz | 614.4 KB | 表示 | |
| CIF形式データ | 4v5m_validation.cif.gz | 854.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/v5/4v5mftp://data.pdbj.org/pub/pdb/validation_reports/v5/4v5m | HTTPS FTP |
-関連構造データ
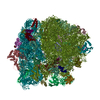
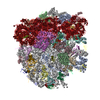
| 関連構造データ | 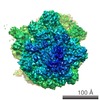 1798MC  1799C 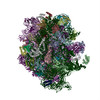 4v5nC  2jl5 2jl7 M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |















-リンク
 PDBj
PDBj





























- 集合体
集合体
| 登録構造単位 | 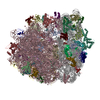
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 5種, 5分子 AAAVAXBABB
| #1: RNA鎖 | 分子量: 493958.281 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: GenBank: 55979969 |
|---|---|
| #22: RNA鎖 | 分子量: 24802.785 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: TRNA WAS COPURIFIED WITH 70S RIBOSOMES / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 |
| #23: RNA鎖 | 分子量: 3476.122 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: MRNA WAS COPURIFIED WITH 70S RIBOSOMES / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 |
| #35: RNA鎖 | 分子量: 947935.438 Da / 分子数: 1 / 由来タイプ: 天然 詳細: CHAIN A (23S RNA) HAS E.COLI RESIDUE NUMBERING, BASED ON A STRUCTURAL ALIGNMENT WITH THE CORRESPONDING E. COLI STRUCTURE IN 2AW4 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 - MRC - MSAW1 / 参照: GenBank: 55979969 |
| #36: RNA鎖 | 分子量: 39540.617 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 - MRC - MSAW1 / 参照: GenBank: 55979969 |
-30S RIBOSOMAL PROTEIN ... , 20種, 20分子 ABACADAEAFAGAHAIAJAKALAMANAOAPAQARASATAU
| #2: タンパク質 | 分子量: 29317.703 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80371 |
|---|---|
| #3: タンパク質 | 分子量: 26751.076 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80372 |
| #4: タンパク質 | 分子量: 24373.447 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80373 |
| #5: タンパク質 | 分子量: 17583.416 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHQ5 |
| #6: タンパク質 | 分子量: 11988.753 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SLP8 |
| #7: タンパク質 | 分子量: 18050.973 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P17291 |
| #8: タンパク質 | 分子量: 15868.570 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHQ2, UniProt: A0A0M9AFS9*PLUS |
| #9: タンパク質 | 分子量: 14410.614 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80374 |
| #10: タンパク質 | 分子量: 11954.968 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHN7 |
| #11: タンパク質 | 分子量: 13737.868 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80376 |
| #12: タンパク質 | 分子量: 14637.384 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHN3 |
| #13: タンパク質 | 分子量: 14338.861 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80377 |
| #14: タンパク質 | 分子量: 7158.725 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHQ1, UniProt: A0A0N0BLP2*PLUS |
| #15: タンパク質 | 分子量: 10578.407 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SJ76 |
| #16: タンパク質 | 分子量: 10409.983 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SJH3 |
| #17: タンパク質 | 分子量: 12325.655 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHP7, UniProt: A0A0N0BLS5*PLUS |
| #18: タンパク質 | 分子量: 10258.299 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SLQ0 |
| #19: タンパク質 | 分子量: 10605.464 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SHP2 |
| #20: タンパク質 | 分子量: 11736.143 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: P80380 |
| #21: タンパク質・ペプチド | 分子量: 3350.030 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) THERMUS THERMOPHILUS (バクテリア) / 株: HB8 / 参照: UniProt: Q5SIH3 |
-タンパク質 , 1種, 1分子 AY
| #24: タンパク質 | 分子量: 76977.102 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) THERMUS THERMOPHILUS (バクテリア)株: HB8 / プラスミド: PET-46 EK/LIC / 発現宿主: |
|---|
+50S RIBOSOMAL PROTEIN ... , 31種, 31分子 B0B1B2B3B4B5B6B7B8B9BCBDBEBFBGBHBKBLBNBOBPBQBRBSBTBUBVBWBXBYBZ
-非ポリマー , 2種, 2分子 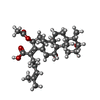
| #58: 化合物 | ChemComp-FUA /  分子量: 516.709 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C31H48O6 / コメント: 抗生剤, 抗菌剤*YM 分子量: 516.709 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C31H48O6 / コメント: 抗生剤, 抗菌剤*YM |
|---|---|
| #59: 化合物 | ChemComp-GDP /  タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM |
-詳細
| 配列の詳細 | MRNA WAS COPURIFIED |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: 70S-EFG-GDP-FA COMPLEX / タイプ: RIBOSOME |
|---|---|
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: HOLEY CARBON |
| 急速凍結 | 装置: FEI VITROBOT MARK I / 凍結剤: ETHANE 詳細: VITRIFICATION 1 -- CRYOGEN- ETHANE, INSTRUMENT- VITROBOT (FEI) |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI POLARA 300 / 詳細: LOW-DOSE |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 39000 X / 倍率(補正後): 65520 X / 最大 デフォーカス(公称値): 4000 nm / 最小 デフォーカス(公称値): 2000 nm / Cs: 2 mm |
| 試料ホルダ | 温度: 77 K |
| 撮影 | 電子線照射量: 20 e/Å2 / フィルム・検出器のモデル: KODAK SO-163 FILM |
| 画像スキャン | デジタル画像の数: 677 |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: DEFOCUS GROUPS | ||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||
| 3次元再構成 | 手法: MULTIPARTICLE REFINEMENT / 解像度: 7.8 Å / ピクセルサイズ(公称値): 1.26 Å / ピクセルサイズ(実測値): 1.26 Å 詳細: SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-1798. 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: OTHER / 空間: REAL / 詳細: METHOD--MDFIT REFINEMENT PROTOCOL--X-RAY | ||||||||||||
| 原子モデル構築 |
| ||||||||||||
| 精密化 | 最高解像度: 7.8 Å | ||||||||||||
| 精密化ステップ | サイクル: LAST / 最高解像度: 7.8 Å
|
