 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
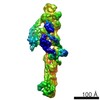
Yorodumi- PDB-6f3a: Cryo-EM structure of a single dynein tail domain bound to dynacti... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6f3a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of a single dynein tail domain bound to dynactin and BICD2N | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | MOTOR PROTEIN / TDB / dynein/dynactin/BICD2 / complex | |||||||||
| Function / homology |  Function and homology information Function and homology information: / Factors involved in megakaryocyte development and platelet production / retrograde axonal transport of mitochondrion / Regulation of actin dynamics for phagocytic cup formation / EPHB-mediated forward signaling / Adherens junctions interactions / VEGFA-VEGFR2 Pathway / Cell-extracellular matrix interactions / RHO GTPases Activate WASPs and WAVEs / MAP2K and MAPK activation ...: / Factors involved in megakaryocyte development and platelet production / retrograde axonal transport of mitochondrion / Regulation of actin dynamics for phagocytic cup formation / EPHB-mediated forward signaling / Adherens junctions interactions / VEGFA-VEGFR2 Pathway / Cell-extracellular matrix interactions / RHO GTPases Activate WASPs and WAVEs / MAP2K and MAPK activation / Formation of the canonical BAF (cBAF) complex / Formation of the polybromo-BAF (pBAF) complex / Formation of the embryonic stem cell BAF (esBAF) complex / Formation of the non-canonical BAF (ncBAF) complex / UCH proteinases / RHOF GTPase cycle / Regulation of CDH1 Function / Formation of the dystrophin-glycoprotein complex (DGC) / Gap junction degradation / Formation of annular gap junctions / Clathrin-mediated endocytosis / dynactin complex / visual behavior / Regulation of PLK1 Activity at G2/M Transition / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Anchoring of the basal body to the plasma membrane / AURKA Activation by TPX2 / Recruitment of mitotic centrosome proteins and complexes / F-actin capping protein complex / WASH complex / dynein light chain binding / transport along microtubule / dynein heavy chain binding / dynein complex / cellular response to cytochalasin B / Intraflagellar transport / positive regulation of intracellular transport / regulation of transepithelial transport / regulation of metaphase plate congression / positive regulation of spindle assembly / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / protein localization to adherens junction / establishment of spindle localization / barbed-end actin filament capping / dense body / Neutrophil degranulation / Tat protein binding / coronary vasculature development / postsynaptic actin cytoskeleton / regulation of cell morphogenesis / adherens junction assembly / retrograde axonal transport / COPI-independent Golgi-to-ER retrograde traffic / apical protein localization / RHO GTPases activate IQGAPs / RHO GTPases Activate Formins / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / minus-end-directed microtubule motor activity / centrosome localization / P-body assembly / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / dynein light intermediate chain binding / cytoplasmic dynein complex / microtubule motor activity / tight junction / ventricular septum development / COPI-mediated anterograde transport / aorta development / microtubule-based movement / nuclear migration / apical junction complex / regulation of norepinephrine uptake / transporter regulator activity / dynein intermediate chain binding / NuA4 histone acetyltransferase complex / cortical cytoskeleton / establishment or maintenance of cell polarity / dynein complex binding / nitric-oxide synthase binding / brush border / regulation of synaptic vesicle endocytosis / kinesin binding / regulation of protein localization to plasma membrane / positive regulation of double-strand break repair via homologous recombination / COPI-mediated anterograde transport / cytoplasmic microtubule / cytoplasmic microtubule organization / cytoskeleton organization / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / axon cytoplasm / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Recruitment of mitotic centrosome proteins and complexes / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / axonogenesis / Anchoring of the basal body to the plasma membrane Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 8.2 Å | |||||||||
 Authors Authors | Urnavicius, L. / Lau, C.K. / Elshenawy, M.M. / Morales-Rios, E. / Motz, C. / Yildiz, A. / Carter, A.P. | |||||||||
| Funding support |  United Kingdom, 2items United Kingdom, 2items
| |||||||||
 Citation Citation | Journal: Nature / Year: 2018 Title: Cryo-EM shows how dynactin recruits two dyneins for faster movement. Authors: Linas Urnavicius / Clinton K Lau / Mohamed M Elshenawy / Edgar Morales-Rios / Carina Motz / Ahmet Yildiz / Andrew P Carter /   Abstract: Dynein and its cofactor dynactin form a highly processive microtubule motor in the presence of an activating adaptor, such as BICD2. Different adaptors link dynein and dynactin to distinct cargoes. ...Dynein and its cofactor dynactin form a highly processive microtubule motor in the presence of an activating adaptor, such as BICD2. Different adaptors link dynein and dynactin to distinct cargoes. Here we use electron microscopy and single-molecule studies to show that adaptors can recruit a second dynein to dynactin. Whereas BICD2 is biased towards recruiting a single dynein, the adaptors BICDR1 and HOOK3 predominantly recruit two dyneins. We find that the shift towards a double dynein complex increases both the force and speed of the microtubule motor. Our 3.5 Å resolution cryo-electron microscopy reconstruction of a dynein tail-dynactin-BICDR1 complex reveals how dynactin can act as a scaffold to coordinate two dyneins side-by-side. Our work provides a structural basis for understanding how diverse adaptors recruit different numbers of dyneins and regulate the motile properties of the dynein-dynactin transport machine. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6f3a.cif.gz | 1.3 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6f3a.ent.gz | 937.4 KB | Display | PDB format |
| PDBx/mmJSON format | 6f3a.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/f3/6f3aftp://data.pdbj.org/pub/pdb/validation_reports/f3/6f3a | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 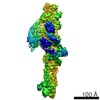 2860M  4168C  4169C  4170C  4171C  4172C  4177C  5owoC  6f1tC  6f1uC  6f1vC  6f1yC  6f1zC  6f38C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj




















- Assembly
Assembly
| Deposited unit | 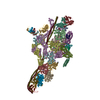
|
|---|---|
| 1 |
|
-Components
-Protein , 8 types, 17 molecules ABCDEFGIHJKLUkl56
| #1: Protein | Mass: 42670.688 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Details: ADP: Adenosine diphosphate / Source: (natural) #2: Protein | | Mass: 41782.660 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Details: ATP: Adenosine triphosphate / Source: (natural) #3: Protein | | Mass: 43203.250 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Details: ADP: Adenosine diphosphate / Source: (natural) #4: Protein | | Mass: 33059.848 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #5: Protein | | Mass: 30669.768 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #10: Protein | | Mass: 20703.910 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #19: Protein | Mass: 10934.576 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNLRB1, BITH, DNCL2A, DNLC2A, ROBLD1, HSPC162 / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9NP97 Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9NP97#21: Protein | Mass: 23421.752 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Spodoptera frugiperda (fall armyworm) |
|---|
-Dynactin Subunit ... , 10 types, 14 molecules MNOPQRVYZabcdz
| #6: Protein | Mass: 49974.707 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| #7: Protein | Mass: 52442.777 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||||||||||||||
| #8: Protein | Mass: 5549.833 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #9: Protein | Mass: 7422.140 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #11: Protein | | Mass: 20150.533 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #12: Protein | | Mass: 22485.607 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #13: Protein | | Mass: 4443.468 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #14: Protein | | Mass: 7578.217 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #15: Protein | Mass: 10015.838 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) #20: Protein | | Mass: 4528.573 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Cytoplasmic dynein 1 ... , 3 types, 5 molecules efghj
| #16: Protein | Mass: 168318.781 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1H1, DHC1, DNCH1, DNCL, DNECL, DYHC, KIAA0325 / Cell line (production host): Sf9 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q14204#17: Protein | Mass: 68442.141 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1I2, DNCI2, DNCIC2 / Cell line (production host): Sf9 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q13409#18: Protein | | Mass: 54173.156 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1LI2, DNCLI2, LIC2 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: O43237 |
|---|
-Non-polymers , 2 types, 10 molecules 

| #22: Chemical | ChemComp-ADP /  Mass: 427.201 Da / Num. of mol.: 9 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 9 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM#23: Chemical | ChemComp-ATP / |  Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM |
|---|
-Details
| Has protein modification | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source (natural) |
| ||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.4 | ||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 51 e/Å2 / Film or detector model: FEI FALCON II (4k x 4k) |
- Processing
Processing
| Software | Name: PHENIX / Version: dev_2919: / Classification: refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 8.2 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 85744 / Symmetry type: POINT | ||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: Cross-correlation coefficient / Details: Took components from PDB 6F1T and fit into map | ||||||||||||||||||||||||
| Refine LS restraints |
|
