 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5owo: Human cytoplasmic Dynein N-Terminus dimerization domain at 1.8 An... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5owo | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Human cytoplasmic Dynein N-Terminus dimerization domain at 1.8 Angstrom resolution | ||||||||||||||||||
 Components Components | Cytoplasmic dynein 1 heavy chain 1 | ||||||||||||||||||
 Keywords Keywords | MOTOR PROTEIN / Dynein / Heavy chain / Dimerization domain | ||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of intracellular transport / regulation of metaphase plate congression / positive regulation of spindle assembly / establishment of spindle localization / retrograde axonal transport / COPI-independent Golgi-to-ER retrograde traffic / minus-end-directed microtubule motor activity / P-body assembly / dynein light intermediate chain binding / cytoplasmic dynein complex ...positive regulation of intracellular transport / regulation of metaphase plate congression / positive regulation of spindle assembly / establishment of spindle localization / retrograde axonal transport / COPI-independent Golgi-to-ER retrograde traffic / minus-end-directed microtubule motor activity / P-body assembly / dynein light intermediate chain binding / cytoplasmic dynein complex / nuclear migration / dynein intermediate chain binding / COPI-mediated anterograde transport / cytoplasmic microtubule / cytoplasmic microtubule organization / axon cytoplasm / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Recruitment of mitotic centrosome proteins and complexes / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / AURKA Activation by TPX2 / Resolution of Sister Chromatid Cohesion / stress granule assembly / mitotic spindle organization / regulation of mitotic spindle organization / filopodium / RHO GTPases Activate Formins / HCMV Early Events / Aggrephagy / azurophil granule lumen / Separation of Sister Chromatids / Regulation of PLK1 Activity at G2/M Transition / positive regulation of cold-induced thermogenesis / cell cortex / microtubule / cell division / Neutrophil degranulation / centrosome / RNA binding / extracellular exosome / extracellular region / ATP binding / membrane / identical protein binding / cytosol Similarity search - Function | ||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MAD / Resolution: 1.79 Å X-RAY DIFFRACTION / SYNCHROTRON / MAD / Resolution: 1.79 Å | ||||||||||||||||||
 Authors Authors | Urnavicius, L. / Lau, C.K. / Elshenawy, M.M. / Morales-Rios, E. / Motz, C. / Yildiz, A. / Carter, A.P. | ||||||||||||||||||
| Funding support |  United Kingdom, United Kingdom,  United States, 5items United States, 5items
| ||||||||||||||||||
 Citation Citation | Journal: Nature / Year: 2018 Title: Cryo-EM shows how dynactin recruits two dyneins for faster movement. Authors: Linas Urnavicius / Clinton K Lau / Mohamed M Elshenawy / Edgar Morales-Rios / Carina Motz / Ahmet Yildiz / Andrew P Carter /  Abstract: Dynein and its cofactor dynactin form a highly processive microtubule motor in the presence of an activating adaptor, such as BICD2. Different adaptors link dynein and dynactin to distinct cargoes. ...Dynein and its cofactor dynactin form a highly processive microtubule motor in the presence of an activating adaptor, such as BICD2. Different adaptors link dynein and dynactin to distinct cargoes. Here we use electron microscopy and single-molecule studies to show that adaptors can recruit a second dynein to dynactin. Whereas BICD2 is biased towards recruiting a single dynein, the adaptors BICDR1 and HOOK3 predominantly recruit two dyneins. We find that the shift towards a double dynein complex increases both the force and speed of the microtubule motor. Our 3.5 Å resolution cryo-electron microscopy reconstruction of a dynein tail-dynactin-BICDR1 complex reveals how dynactin can act as a scaffold to coordinate two dyneins side-by-side. Our work provides a structural basis for understanding how diverse adaptors recruit different numbers of dyneins and regulate the motile properties of the dynein-dynactin transport machine. | ||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5owo.cif.gz | 147.4 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5owo.ent.gz | 116.4 KB | Display | PDB format |
| PDBx/mmJSON format | 5owo.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ow/5owoftp://data.pdbj.org/pub/pdb/validation_reports/ow/5owo | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4168C  4169C  4170C  4171C  4172C  4177C  6f1tC  6f1uC  6f1vC  6f1yC 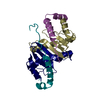 6f1zC  6f38C  6f3aC C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj










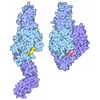





- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| Unit cell |
| ||||||||||||
| Components on special symmetry positions |
|
-Components
-Protein , 1 types, 4 molecules ABCD
| #1: Protein | Mass: 22147.223 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Details: Selenomethionine labeled protein. / Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1H1, DHC1, DNCH1, DNCL, DNECL, DYHC, KIAA0325 / Production host:  |
|---|
-Non-polymers , 6 types, 313 molecules 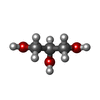





| #2: Chemical | ChemComp-GOL /  Mass: 92.094 Da / Num. of mol.: 9 / Source method: obtained synthetically / Formula: C3H8O3 Mass: 92.094 Da / Num. of mol.: 9 / Source method: obtained synthetically / Formula: C3H8O3#3: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 26 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 26 / Source method: obtained synthetically / Formula: Mg#4: Chemical |  Mass: 40.078 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Ca Mass: 40.078 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Ca#5: Chemical | ChemComp-NA / |  Mass: 22.990 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Na Mass: 22.990 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Na#6: Chemical | ChemComp-K / |  Mass: 39.098 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: K Mass: 39.098 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: K#7: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 274 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.55 Å3/Da / Density % sol: 51.8 % |
|---|---|
| Crystal grow | Temperature: 291.15 K / Method: vapor diffusion, hanging drop / pH: 5.5 Details: 100mM Sodium acetate, pH 5.5, 10% Glycerol, 50mM Calcium acetate, 20% PEG 2,000 MME |
-Data collection
| Diffraction | Mean temperature: 100 K | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: ESRF  / Beamline: ID29 / Wavelength: 0.9793, 0.9792 / Beamline: ID29 / Wavelength: 0.9793, 0.9792 | |||||||||
| Detector | Type: DECTRIS PILATUS3 6M / Detector: PIXEL / Date: Feb 6, 2016 | |||||||||
| Radiation | Protocol: MAD / Monochromatic (M) / Laue (L): M / Scattering type: x-ray | |||||||||
| Radiation wavelength |
| |||||||||
| Reflection | Resolution: 1.6→44.29 Å / Num. obs: 119216 / % possible obs: 98.5 % / Redundancy: 3.07 % / Biso Wilson estimate: 25.7 Å2 / Rmerge(I) obs: 0.2 / Net I/σ(I): 3.9 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MAD / Resolution: 1.79→87.78 Å / Cor.coef. Fo:Fc: 0.966 / Cor.coef. Fo:Fc free: 0.954 / SU B: 3.18 / SU ML: 0.092 / Cross valid method: THROUGHOUT / ESU R: 0.109 / ESU R Free: 0.109 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 31.823 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: 1 / Resolution: 1.79→87.78 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|