 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7by0 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
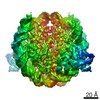
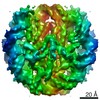
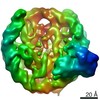
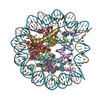
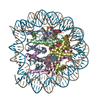
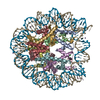
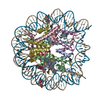
| タイトル | The cryo-EM structure of CENP-A nucleosome in complex with the phosphorylated CENP-C | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | CELL CYCLE / CENP-A nucleosome / CENP-C / CENP-N / complex / kinetochore / NUCLEAR PROTEIN | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Interleukin-7 signaling / Chromatin modifying enzymes / Formation of the beta-catenin:TCF transactivating complex / PRC2 methylates histones and DNA / Oxidative Stress Induced Senescence / HDACs deacetylate histones / HATs acetylate histones / Transcriptional regulation by small RNAs / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / Assembly of the ORC complex at the origin of replication ...Interleukin-7 signaling / Chromatin modifying enzymes / Formation of the beta-catenin:TCF transactivating complex / PRC2 methylates histones and DNA / Oxidative Stress Induced Senescence / HDACs deacetylate histones / HATs acetylate histones / Transcriptional regulation by small RNAs / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / Assembly of the ORC complex at the origin of replication / RNA Polymerase I Promoter Opening / RNA Polymerase I Promoter Escape / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Estrogen-dependent gene expression / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / Regulation of endogenous retroelements by KRAB-ZFP proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / B-WICH complex positively regulates rRNA expression / Factors involved in megakaryocyte development and platelet production / CENP-A containing chromatin assembly / centromeric DNA binding / kinetochore assembly / protein localization to chromosome, centromeric region / condensed chromosome, centromeric region / detection of maltose stimulus / maltose transport complex / carbohydrate transport / mitotic cytokinesis / establishment of mitotic spindle orientation / carbohydrate transmembrane transporter activity / maltose binding / negative regulation of tumor necrosis factor-mediated signaling pathway / maltose transport / maltodextrin transmembrane transport / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / ATP-binding cassette (ABC) transporter complex, substrate-binding subunit-containing / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / telomere organization / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / ATP-binding cassette (ABC) transporter complex / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / innate immune response in mucosa / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / cell chemotaxis / HDMs demethylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / Nonhomologous End-Joining (NHEJ) / lipopolysaccharide binding / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / G2/M DNA damage checkpoint / Metalloprotease DUBs / NoRC negatively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / B-WICH complex positively regulates rRNA expression / kinetochore / PKMTs methylate histone lysines / Meiotic recombination / Pre-NOTCH Transcription and Translation / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / UCH proteinases / HCMV Early Events / antimicrobial humoral immune response mediated by antimicrobial peptide / structural constituent of chromatin / antibacterial humoral response / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / nucleosome / heterochromatin formation / RUNX1 regulates transcription of genes involved in differentiation of HSCs / nucleosome assembly / Processing of DNA double-strand break ends / HATs acetylate histones / Senescence-Associated Secretory Phenotype (SASP) 類似検索 - 分子機能 | ||||||||||||
| 生物種 |   Homo sapiens (ヒト) Homo sapiens (ヒト)unidentified cloning vector (未定義) synthetic construct (人工物) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.5 Å | ||||||||||||
 データ登録者 データ登録者 | Ariyoshi, M. / Makino, F. / Fukagawa, T. | ||||||||||||
| 資金援助 |  日本, 3件 日本, 3件
| ||||||||||||
 引用 引用 | ジャーナル: EMBO J / 年: 2021 タイトル: Cryo-EM structure of the CENP-A nucleosome in complex with phosphorylated CENP-C. 著者: Mariko Ariyoshi / Fumiaki Makino / Reito Watanabe / Reiko Nakagawa / Takayuki Kato / Keiichi Namba / Yasuhiro Arimura / Risa Fujita / Hitoshi Kurumizaka / Ei-Ichi Okumura / Masatoshi Hara / Tatsuo Fukagawa / 要旨: The CENP-A nucleosome is a key structure for kinetochore assembly. Once the CENP-A nucleosome is established in the centromere, additional proteins recognize the CENP-A nucleosome to form a ...The CENP-A nucleosome is a key structure for kinetochore assembly. Once the CENP-A nucleosome is established in the centromere, additional proteins recognize the CENP-A nucleosome to form a kinetochore. CENP-C and CENP-N are CENP-A binding proteins. We previously demonstrated that vertebrate CENP-C binding to the CENP-A nucleosome is regulated by CDK1-mediated CENP-C phosphorylation. However, it is still unknown how the phosphorylation of CENP-C regulates its binding to CENP-A. It is also not completely understood how and whether CENP-C and CENP-N act together on the CENP-A nucleosome. Here, using cryo-electron microscopy (cryo-EM) in combination with biochemical approaches, we reveal a stable CENP-A nucleosome-binding mode of CENP-C through unique regions. The chicken CENP-C structure bound to the CENP-A nucleosome is stabilized by an intramolecular link through the phosphorylated CENP-C residue. The stable CENP-A-CENP-C complex excludes CENP-N from the CENP-A nucleosome. These findings provide mechanistic insights into the dynamic kinetochore assembly regulated by CDK1-mediated CENP-C phosphorylation. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7by0.cif.gz | 327.9 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7by0.ent.gz | 233.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7by0.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7by0_validation.pdf.gz | 976 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7by0_full_validation.pdf.gz | 997 KB | 表示 | |
| XML形式データ | 7by0_validation.xml.gz | 38.7 KB | 表示 | |
| CIF形式データ | 7by0_validation.cif.gz | 60.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/by/7by0ftp://data.pdbj.org/pub/pdb/validation_reports/by/7by0 | HTTPS FTP |
-関連構造データ
| 関連構造データ | 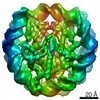 30239MC  7bxtC M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ | |
| 電子顕微鏡画像生データ | EMPIAR-10565 (タイトル: The cryo-EM structure of the CENP-A nucleosome in complex with the phosphorylated CENP-C: CENP-A nucleosome in complex with phosphorylated CENP-C C-terminal domain (601-864) and CENP-N ...タイトル: The cryo-EM structure of the CENP-A nucleosome in complex with the phosphorylated CENP-C: CENP-A nucleosome in complex with phosphorylated CENP-C C-terminal domain (601-864) and CENP-N N-terminal domain (1-211) Data size: 1.6 TB Data #1: CENP-A nucleosome in complex with phosphorylated CENP-C C-terminal domain (601-864) and CENP-N N-terminal domain (1-211) [micrographs - multiframe]) |
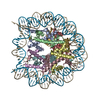

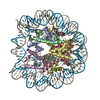
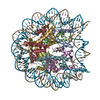
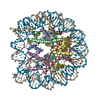

-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 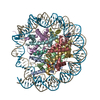
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 10分子 AEBFCGDHKL
| #1: タンパク質 | 分子量: 16089.938 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: chimeric protein of chicken Histone H3.2 (RESIDUES 1-64) and chicken CENP-C (RESIDUES 65-141, UNP RESIDUES 55-131) 由来: (組換発現)  #2: タンパク質 | 分子量: 11337.374 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / プラスミド: pET15b / 発現宿主: #3: タンパク質 | 分子量: 14165.551 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / プラスミド: pET15b / 発現宿主: #4: タンパク質 | 分子量: 13935.239 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HIST1H2BJ, H2BFR / プラスミド: pET15b / 発現宿主: #7: タンパク質 | 分子量: 73796.312 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: fusion protein of N-terminal MBP (RESIDUES 201-600), linkers, and the phosphorylated C-terminal chicken CNPE-C (RESIDUES 612-864, UNP RESIDUES 601-853) 由来: (組換発現) unidentified cloning vector (未定義), (組換発現) Variant: cloning vector pMal-C5X / プラスミド: pMAL_c2X / 発現宿主: |
|---|
-DNA鎖 , 2種, 2分子 IJ
| #5: DNA鎖 | 分子量: 44520.383 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #6: DNA鎖 | 分子量: 44991.660 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.33 MDa / 実験値: NO | ||||||||||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.5 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: MOLYBDENUM / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R0.6/1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 顕微鏡 | モデル: JEOL CRYO ARM 200 |
|---|---|
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 50000 X / 倍率(補正後): 45454 X / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 1000 nm / Calibrated defocus min: -500 nm / 最大 デフォーカス(補正後): -7000 nm / Cs: 1.4 mm / C2レンズ絞り径: 150 µm / アライメント法: BASIC |
| 試料ホルダ | 凍結剤: NITROGEN / 試料ホルダーモデル: JEOL CRYOSPECPORTER / 最高温度: 80 K / 最低温度: 80 K |
| 撮影 | 平均露光時間: 10 sec. / 電子線照射量: 1.2 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 5 / 実像数: 5346 |
| 画像スキャン | サンプリングサイズ: 5 µm / 横: 3838 / 縦: 3710 / 動画フレーム数/画像: 50 / 利用したフレーム数/画像: 1-50 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 343497 | ||||||||||||||||||||||||
| 対称性 | 点対称性: C2 (2回回転対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.5 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 38542 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE |
