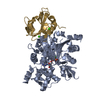
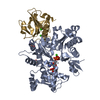
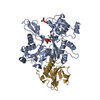
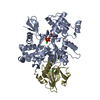
ジャーナル: PLoS Pathog / 年: 2014 タイトル: Structural differences explain diverse functions of Plasmodium actins. 著者: Juha Vahokoski / Saligram Prabhakar Bhargav / Ambroise Desfosses / Maria Andreadaki / Esa-Pekka Kumpula / Silvia Muñico Martinez / Alexander Ignatev / Simone Lepper / Friedrich Frischknecht ...著者: Juha Vahokoski / Saligram Prabhakar Bhargav / Ambroise Desfosses / Maria Andreadaki / Esa-Pekka Kumpula / Silvia Muñico Martinez / Alexander Ignatev / Simone Lepper / Friedrich Frischknecht / Inga Sidén-Kiamos / Carsten Sachse / Inari Kursula / 要旨: Actins are highly conserved proteins and key players in central processes in all eukaryotic cells. The two actins of the malaria parasite are among the most divergent eukaryotic actins and also ...Actins are highly conserved proteins and key players in central processes in all eukaryotic cells. The two actins of the malaria parasite are among the most divergent eukaryotic actins and also differ from each other more than isoforms in any other species. Microfilaments have not been directly observed in Plasmodium and are presumed to be short and highly dynamic. We show that actin I cannot complement actin II in male gametogenesis, suggesting critical structural differences. Cryo-EM reveals that Plasmodium actin I has a unique filament structure, whereas actin II filaments resemble canonical F-actin. Both Plasmodium actins hydrolyze ATP more efficiently than α-actin, and unlike any other actin, both parasite actins rapidly form short oligomers induced by ADP. Crystal structures of both isoforms pinpoint several structural changes in the monomers causing the unique polymerization properties. Inserting the canonical D-loop to Plasmodium actin I leads to the formation of long filaments in vitro. In vivo, this chimera restores gametogenesis in parasites lacking actin II, suggesting that stable filaments are required for exflagellation. Together, these data underline the divergence of eukaryotic actins and demonstrate how structural differences in the monomers translate into filaments with different properties, implying that even eukaryotic actins have faced different evolutionary pressures and followed different paths for developing their polymerization properties.
履歴
登録
2013年10月17日
登録サイト: PDBE / 処理サイト: PDBE
改定 1.0
2014年4月30日
Provider: repository / タイプ: Initial release
改定 1.1
2014年5月14日
Group: Data collection
改定 1.2
2017年3月15日
Group: Source and taxonomy
改定 2.0
2019年10月23日
Group: Atomic model / Data collection / Other / カテゴリ: atom_site / pdbx_database_status Item: _atom_site.B_iso_or_equiv / _atom_site.Cartn_x ..._atom_site.B_iso_or_equiv / _atom_site.Cartn_x / _atom_site.Cartn_y / _atom_site.Cartn_z / _pdbx_database_status.status_code_sf
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報

 X線回折 /
X線回折 /  データ登録者
データ登録者 引用
引用


 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード






 PDBj
PDBj










 集合体
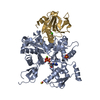
集合体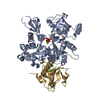

 SPODOPTERA FRUGIPERDA (ツマジロクサヨトウ)
SPODOPTERA FRUGIPERDA (ツマジロクサヨトウ)



 分子量: 40.078 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Ca
分子量: 40.078 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Ca 分子量: 88.105 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C4H8O2
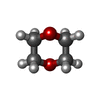
分子量: 88.105 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C4H8O2 分子量: 507.181 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / コメント: ATP, エネルギー貯蔵分子*YM
分子量: 507.181 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / コメント: ATP, エネルギー貯蔵分子*YM 試料調製
試料調製 解析
解析