 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4cbw: Crystal structure of Plasmodium berghei actin I with D-loop from ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4cbw | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of Plasmodium berghei actin I with D-loop from muscle actin | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | MOTOR PROTEIN / MALARIA / MOTILITY / PARASITE | |||||||||
| Function / homology |  Function and homology information Function and homology informationglial filament / basal ectoplasmic specialization / apical ectoplasmic specialization / Caspase-mediated cleavage of cytoskeletal proteins / regulation of podosome assembly / striated muscle atrophy / regulation of establishment of T cell polarity / regulation of plasma membrane raft polarization / regulation of receptor clustering / positive regulation of keratinocyte apoptotic process ...glial filament / basal ectoplasmic specialization / apical ectoplasmic specialization / Caspase-mediated cleavage of cytoskeletal proteins / regulation of podosome assembly / striated muscle atrophy / regulation of establishment of T cell polarity / regulation of plasma membrane raft polarization / regulation of receptor clustering / positive regulation of keratinocyte apoptotic process / renal protein absorption / positive regulation of protein processing in phagocytic vesicle / phosphatidylinositol 3-kinase catalytic subunit binding / positive regulation of actin nucleation / actin cap / myosin II binding / host-mediated suppression of symbiont invasion / cell projection assembly / actin filament severing / barbed-end actin filament capping / actin polymerization or depolymerization / actin filament depolymerization / actin filament capping / positive regulation of p38MAPK cascade / podosome / relaxation of cardiac muscle / phagocytosis, engulfment / cardiac muscle cell contraction / hepatocyte apoptotic process / cortical actin cytoskeleton / positive regulation of cardiac muscle hypertrophy / cellular response to cadmium ion / sarcoplasm / cilium assembly / phagocytic vesicle / ruffle / response to muscle stretch / vesicle-mediated transport / actin filament polymerization / Neutrophil degranulation / phosphatidylinositol-4,5-bisphosphate binding / actin filament organization / central nervous system development / cellular response to type II interferon / protein destabilization / Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement / actin filament binding / myelin sheath / lamellipodium / actin cytoskeleton / actin cytoskeleton organization / actin binding / amyloid fibril formation / cytoskeleton / calcium ion binding / positive regulation of gene expression / perinuclear region of cytoplasm / ATP hydrolysis activity / protein-containing complex / : / extracellular region / ATP binding / nucleus / plasma membrane / cytosol / cytoplasm Similarity search - Function | |||||||||
| Biological species |  SYNTHETIC CONSTRUCT (others)  | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.501 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.501 Å | |||||||||
 Authors Authors | Vahokoski, J. / Bhargav, S.P. / Desfosses, A. / Andreadaki, M. / Kumpula, E.P. / Ignatev, A. / Munico Martinez, S. / Lepper, S. / Frischknecht, F. / Siden-Kiamos, I. ...Vahokoski, J. / Bhargav, S.P. / Desfosses, A. / Andreadaki, M. / Kumpula, E.P. / Ignatev, A. / Munico Martinez, S. / Lepper, S. / Frischknecht, F. / Siden-Kiamos, I. / Sachse, C. / Kursula, I. | |||||||||
 Citation Citation | Journal: PLoS Pathog / Year: 2014 Title: Structural differences explain diverse functions of Plasmodium actins. Authors: Juha Vahokoski / Saligram Prabhakar Bhargav / Ambroise Desfosses / Maria Andreadaki / Esa-Pekka Kumpula / Silvia Muñico Martinez / Alexander Ignatev / Simone Lepper / Friedrich Frischknecht ...Authors: Juha Vahokoski / Saligram Prabhakar Bhargav / Ambroise Desfosses / Maria Andreadaki / Esa-Pekka Kumpula / Silvia Muñico Martinez / Alexander Ignatev / Simone Lepper / Friedrich Frischknecht / Inga Sidén-Kiamos / Carsten Sachse / Inari Kursula /    Abstract: Actins are highly conserved proteins and key players in central processes in all eukaryotic cells. The two actins of the malaria parasite are among the most divergent eukaryotic actins and also ...Actins are highly conserved proteins and key players in central processes in all eukaryotic cells. The two actins of the malaria parasite are among the most divergent eukaryotic actins and also differ from each other more than isoforms in any other species. Microfilaments have not been directly observed in Plasmodium and are presumed to be short and highly dynamic. We show that actin I cannot complement actin II in male gametogenesis, suggesting critical structural differences. Cryo-EM reveals that Plasmodium actin I has a unique filament structure, whereas actin II filaments resemble canonical F-actin. Both Plasmodium actins hydrolyze ATP more efficiently than α-actin, and unlike any other actin, both parasite actins rapidly form short oligomers induced by ADP. Crystal structures of both isoforms pinpoint several structural changes in the monomers causing the unique polymerization properties. Inserting the canonical D-loop to Plasmodium actin I leads to the formation of long filaments in vitro. In vivo, this chimera restores gametogenesis in parasites lacking actin II, suggesting that stable filaments are required for exflagellation. Together, these data underline the divergence of eukaryotic actins and demonstrate how structural differences in the monomers translate into filaments with different properties, implying that even eukaryotic actins have faced different evolutionary pressures and followed different paths for developing their polymerization properties. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4cbw.cif.gz | 298.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4cbw.ent.gz | 244.8 KB | Display | PDB format |
| PDBx/mmJSON format | 4cbw.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/cb/4cbwftp://data.pdbj.org/pub/pdb/validation_reports/cb/4cbw | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  2572C  4cbuSC  4cbxC S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj










- Assembly
Assembly
| Deposited unit | 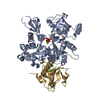
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
-Protein , 2 types, 2 molecules AG
| #1: Protein | Mass: 41944.480 Da / Num. of mol.: 1 Fragment: ACTIN RESIDUES 2-39, ALPHA SKELETAL MUSCLE RESIDUES 40-61, Source method: isolated from a genetically manipulated source Details: RESIDUES 40-61 REPLACED BY THOSE OF SKELETAL MUSCLE ACTIN Source: (gene. exp.) Production host:   SPODOPTERA FRUGIPERDA (fall armyworm) / Strain (production host): SF21 / References: UniProt: Q4Z1L3 SPODOPTERA FRUGIPERDA (fall armyworm) / Strain (production host): SF21 / References: UniProt: Q4Z1L3 |
|---|---|
| #2: Protein | Mass: 14211.929 Da / Num. of mol.: 1 / Fragment: G1 DOMAIN, RESIDUES 25-149 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  |
-Non-polymers , 4 types, 61 molecules 
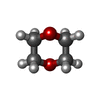


| #3: Chemical |  Mass: 40.078 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Ca Mass: 40.078 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Ca#4: Chemical | ChemComp-DIO / |  Mass: 88.105 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C4H8O2 Mass: 88.105 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C4H8O2#5: Chemical | ChemComp-ATP / |  Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM#6: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 56 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.9 Å3/Da / Density % sol: 58 % / Description: NONE |
|---|---|
| Crystal grow | pH: 8 Details: 100 MM TRIS-HCL (PH 8.0), 8% (W/V), PEG 20000, 2% (V/V) DIOXANE |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: PETRA III, DESY / Beamline: P11 / Wavelength: 0.92 |
| Detector | Type: DECTRIS PILATUS 6M / Detector: PIXEL / Date: Mar 1, 2013 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.92 Å / Relative weight: 1 |
| Reflection | Resolution: 2.5→38.5 Å / Num. obs: 24163 / % possible obs: 99.6 % / Observed criterion σ(I): -1 / Redundancy: 6.7 % / Biso Wilson estimate: 55.19 Å2 / Rmerge(I) obs: 0.16 / Net I/σ(I): 9.2 |
| Reflection shell | Resolution: 2.5→2.6 Å / Redundancy: 4.1 % / Rmerge(I) obs: 1.5 / Mean I/σ(I) obs: 0.9 / % possible all: 96.9 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRY 4CBU Resolution: 2.501→38.581 Å / SU ML: 0.36 / σ(F): 1.37 / Phase error: 32.92 / Stereochemistry target values: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 75 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.501→38.581 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
