ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8txw | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
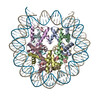
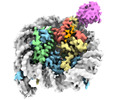
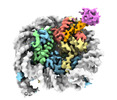
| タイトル | Cryo-EM structure of the human nucleosome core particle ubiquitylated at histone H2A K15 in complex with RNF168 (Class 2) | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | TRANSFERASE / Nucleosome core particle / chromatin / RNF168 / MIU2-LRM domains / DNA repair / DNA double-strand break / Homologous recombination / BRCA1-BARD1 / 53BP1 / ubiquitin / STRUCTURAL PROTEIN-DNA-TRANSFERASE complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報histone H2AK15 ubiquitin ligase activity / histone ubiquitin ligase activity / double-strand break repair via classical nonhomologous end joining / isotype switching / hypothalamus gonadotrophin-releasing hormone neuron development / female meiosis I / positive regulation of protein monoubiquitination / DNA repair-dependent chromatin remodeling / mitochondrion transport along microtubule / fat pad development ...histone H2AK15 ubiquitin ligase activity / histone ubiquitin ligase activity / double-strand break repair via classical nonhomologous end joining / isotype switching / hypothalamus gonadotrophin-releasing hormone neuron development / female meiosis I / positive regulation of protein monoubiquitination / DNA repair-dependent chromatin remodeling / mitochondrion transport along microtubule / fat pad development / K63-linked polyubiquitin modification-dependent protein binding / response to ionizing radiation / female gonad development / seminiferous tubule development / male meiosis I / negative regulation of transcription elongation by RNA polymerase II / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / protein K63-linked ubiquitination / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / interstrand cross-link repair / SUMOylation of DNA damage response and repair proteins / energy homeostasis / regulation of neuron apoptotic process / heterochromatin organization / epigenetic regulation of gene expression / Packaging Of Telomere Ends / regulation of proteasomal protein catabolic process / ubiquitin ligase complex / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / nucleosome binding / Maturation of protein E / Deposition of new CENPA-containing nucleosomes at the centromere / Maturation of protein E / nucleosomal DNA binding / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / Prevention of phagosomal-lysosomal fusion / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Membrane binding and targetting of GAG proteins / Endosomal Sorting Complex Required For Transport (ESCRT) / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Negative regulation of FLT3 / Constitutive Signaling by NOTCH1 HD Domain Mutants / Regulation of FZD by ubiquitination / TICAM1,TRAF6-dependent induction of TAK1 complex / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1-dependent activation of IRF3/IRF7 / Inhibition of DNA recombination at telomere / Meiotic synapsis / APC/C:Cdc20 mediated degradation of Cyclin B / p75NTR recruits signalling complexes / positive regulation of DNA repair / Downregulation of ERBB4 signaling / telomere organization / APC-Cdc20 mediated degradation of Nek2A / PINK1-PRKN Mediated Mitophagy / TRAF6-mediated induction of TAK1 complex within TLR4 complex / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / Pexophagy / Regulation of innate immune responses to cytosolic DNA / InlA-mediated entry of Listeria monocytogenes into host cells / VLDLR internalisation and degradation / RNA Polymerase I Promoter Opening / Interleukin-7 signaling / Downregulation of ERBB2:ERBB3 signaling / NF-kB is activated and signals survival / NRIF signals cell death from the nucleus / Regulation of PTEN localization / Assembly of the ORC complex at the origin of replication / Activated NOTCH1 Transmits Signal to the Nucleus / SUMOylation of chromatin organization proteins / Regulation of BACH1 activity / Translesion synthesis by REV1 / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by POLK / MAP3K8 (TPL2)-dependent MAPK1/3 activation / TICAM1, RIP1-mediated IKK complex recruitment / Downregulation of TGF-beta receptor signaling / DNA methylation / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / Josephin domain DUBs / neuron projection morphogenesis / Regulation of activated PAK-2p34 by proteasome mediated degradation / Condensation of Prophase Chromosomes / InlB-mediated entry of Listeria monocytogenes into host cell 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.6 Å | |||||||||
 データ登録者 データ登録者 | Hu, Q. / Botuyan, M.V. / Zhao, D. / Cui, G. / Mer, G. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2024 タイトル: Mechanisms of RNF168 nucleosome recognition and ubiquitylation. 著者: Qi Hu / Debiao Zhao / Gaofeng Cui / Janarjan Bhandari / James R Thompson / Maria Victoria Botuyan / Georges Mer / 要旨: RNF168 plays a central role in the DNA damage response (DDR) by ubiquitylating histone H2A at K13 and K15. These modifications direct BRCA1-BARD1 and 53BP1 foci formation in chromatin, essential for ...RNF168 plays a central role in the DNA damage response (DDR) by ubiquitylating histone H2A at K13 and K15. These modifications direct BRCA1-BARD1 and 53BP1 foci formation in chromatin, essential for cell-cycle-dependent DNA double-strand break (DSB) repair pathway selection. The mechanism by which RNF168 catalyzes the targeted accumulation of H2A ubiquitin conjugates to form repair foci around DSBs remains unclear. Here, using cryoelectron microscopy (cryo-EM), nuclear magnetic resonance (NMR) spectroscopy, and functional assays, we provide a molecular description of the reaction cycle and dynamics of RNF168 as it modifies the nucleosome and recognizes its ubiquitylation products. We demonstrate an interaction of a canonical ubiquitin-binding domain within full-length RNF168, which not only engages ubiquitin but also the nucleosome surface, clarifying how such site-specific ubiquitin recognition propels a signal amplification loop. Beyond offering mechanistic insights into a key DDR protein, our study aids in understanding site specificity in both generating and interpreting chromatin ubiquitylation. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8txw.cif.gz | 316.9 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8txw.ent.gz | 232.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8txw.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8txw_validation.pdf.gz | 1.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8txw_full_validation.pdf.gz | 1.2 MB | 表示 | |
| XML形式データ | 8txw_validation.xml.gz | 37.1 KB | 表示 | |
| CIF形式データ | 8txw_validation.cif.gz | 57.9 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/tx/8txwftp://data.pdbj.org/pub/pdb/validation_reports/tx/8txw | HTTPS FTP |
-関連構造データ
| 関連構造データ |  41707MC  8smwC  8smxC  8smyC  8smzC  8sn0C  8sn1C  8sn2C 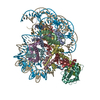 8sn3C  8sn4C  8sn5C  8sn6C  8sn7C  8sn8C  8sn9C  8snaC  8txvC  8txxC  8u13C  8u14C  8upfC  8uq8C  8uq9C  8uqaC  8uqbC  8uqcC  8uqdC  8uqeC M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |
















-リンク
 PDBj
PDBj





















































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 6種, 10分子 AEBFDHKMCG
| #1: タンパク質 | 分子量: 15786.534 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H3C1, H3FA, HIST1H3A, H3C2, H3FL, HIST1H3B, H3C3, H3FC HIST1H3C, H3C4, H3FB, HIST1H3D, H3C6, H3FD, HIST1H3E, H3C7, H3FI, HIST1H3F, H3C8, H3FH, HIST1H3G, H3C10, H3FK, HIST1H3H, H3C11, H3FF, ...遺伝子: H3C1, H3FA, HIST1H3A, H3C2, H3FL, HIST1H3B, H3C3, H3FC HIST1H3C, H3C4, H3FB, HIST1H3D, H3C6, H3FD, HIST1H3E, H3C7, H3FI, HIST1H3F, H3C8, H3FH, HIST1H3G, H3C10, H3FK, HIST1H3H, H3C11, H3FF, HIST1H3I, H3C12, H3FJ, HIST1H3J プラスミド: pHISPP / 発現宿主:  #2: タンパク質 | 分子量: 11743.792 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, ...遺伝子: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, HIST1H4H, H4C9, H4/M, H4FM, HIST1H4I, H4C11, H4/E, H4FE, HIST1H4J, H4C12, H4/D, H4FD, HIST1H4K, H4C13, H4/K, H4FK, HIST1H4L, H4C14, H4/N, H4F2, H4FN, HIST2H4, HIST2H4A, H4C15, H4/O, H4FO, HIST2H4B, H4-16, HIST4H4 プラスミド: pHISPP / 発現宿主: #3: タンパク質 | 分子量: 14084.348 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H2BC4, H2BFL, HIST1H2BC, H2BC6, H2BFH, HIST1H2BE, H2BC7, H2BFG, HIST1H2BF, H2BC8, H2BFA, HIST1H2BG, H2BC10, H2BFK, HIST1H2BI プラスミド: pHISPP / 発現宿主: #6: タンパク質 | | 分子量: 66168.180 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RNF168 / プラスミド: pHISPP / 発現宿主: #7: タンパク質 | | 分子量: 9057.391 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: UBB / プラスミド: pHISPP / 発現宿主: #8: タンパク質 | 分子量: 12992.091 Da / 分子数: 2 / Mutation: K13S / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: H2AC4, H2AFM, HIST1H2AB, H2AC8, H2AFA, HIST1H2AE / プラスミド: pHISPP / 発現宿主: |
|---|
-DNA鎖 , 2種, 2分子 IJ
| #4: DNA鎖 | 分子量: 45138.770 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #5: DNA鎖 | 分子量: 45610.043 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Human nucleosome core particle ubiquitylated at histone H2A K15 in complex with RNF168 (Class 2) タイプ: COMPLEX / Entity ID: all / 由来: MULTIPLE SOURCES |
|---|---|
| 分子量 | 値: 0.28 MDa / 実験値: NO |
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 7.5 |
| 試料 | 濃度: 0.25 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES / 詳細: 10 mM HEPES, 100 mM NaCl, 1 mM DTT, pH 7.5 |
| 試料支持 | グリッドの材料: COPPER / グリッドのタイプ: Quantifoil R1.2/1.3 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 130000 X / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 500 nm / Cs: 2.7 mm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 50 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 11712 詳細: 11712 images were recorded in movie-mode of which 10993 were retained for particle picking. |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 9619981 | ||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 47733 / アルゴリズム: FOURIER SPACE / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
| ||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|