 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7oo3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Pol II-CSB-CSA-DDB1-UVSSA (Structure1) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 要素 要素 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 キーワード キーワード | TRANSCRIPTION / DNA repair | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報RNA polymerase inhibitor activity / regulation of transcription-coupled nucleotide-excision repair / negative regulation of double-strand break repair via nonhomologous end joining / nucleotide-excision repair complex / : / response to auditory stimulus / regulation of transcription elongation by RNA polymerase II / B-WICH complex / DNA protection / single strand break repair ...RNA polymerase inhibitor activity / regulation of transcription-coupled nucleotide-excision repair / negative regulation of double-strand break repair via nonhomologous end joining / nucleotide-excision repair complex / : / response to auditory stimulus / regulation of transcription elongation by RNA polymerase II / B-WICH complex / DNA protection / single strand break repair / Formation of RNA Pol II elongation complex / Formation of the Early Elongation Complex / Transcriptional regulation by small RNAs / RNA Polymerase II Pre-transcription Events / TP53 Regulates Transcription of DNA Repair Genes / FGFR2 alternative splicing / RNA polymerase II transcribes snRNA genes / mRNA Capping / mRNA Splicing - Minor Pathway / Processing of Capped Intron-Containing Pre-mRNA / RNA Polymerase II Promoter Escape / RNA Polymerase II Transcription Pre-Initiation And Promoter Opening / RNA Polymerase II Transcription Initiation / RNA Polymerase II Transcription Elongation / RNA Polymerase II Transcription Initiation And Promoter Clearance / RNA Pol II CTD phosphorylation and interaction with CE / Estrogen-dependent gene expression / response to superoxide / Formation of TC-NER Pre-Incision Complex / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / mRNA Splicing - Major Pathway / double-strand break repair via classical nonhomologous end joining / photoreceptor cell maintenance / chromatin-protein adaptor activity / ATP-dependent chromatin remodeler activity / positive regulation by virus of viral protein levels in host cell / spindle assembly involved in female meiosis / RNA polymerase binding / epigenetic programming in the zygotic pronuclei / positive regulation of DNA-templated transcription, elongation / response to UV-B / UV-damage excision repair / positive regulation of transcription by RNA polymerase III / biological process involved in interaction with symbiont / regulation of mitotic cell cycle phase transition / WD40-repeat domain binding / ATP-dependent DNA damage sensor activity / Cul4A-RING E3 ubiquitin ligase complex / Cul4-RING E3 ubiquitin ligase complex / positive regulation of nuclear-transcribed mRNA poly(A) tail shortening / Cul4B-RING E3 ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / organelle membrane / positive regulation of transcription by RNA polymerase I / negative regulation of reproductive process / negative regulation of developmental process / RNA polymerase II complex binding / protein tyrosine kinase activator activity / RNA Polymerase I Transcription Initiation / maintenance of transcriptional fidelity during transcription elongation by RNA polymerase II / site of DNA damage / viral release from host cell / cullin family protein binding / positive regulation of translational initiation / pyrimidine dimer repair / response to X-ray / positive regulation of transcription initiation by RNA polymerase II / ATP-dependent activity, acting on DNA / ectopic germ cell programmed cell death / RNA polymerase I complex / transcription elongation by RNA polymerase I / RNA polymerase III complex / ubiquitin-like ligase-substrate adaptor activity / positive regulation of viral genome replication / response to UV / RNA polymerase II, core complex / tRNA transcription by RNA polymerase III / positive regulation of double-strand break repair via homologous recombination / protein autoubiquitination / proteasomal protein catabolic process / transcription by RNA polymerase I / JNK cascade / neurogenesis / transcription-coupled nucleotide-excision repair / translation initiation factor binding / positive regulation of gluconeogenesis / DNA damage checkpoint signaling / positive regulation of DNA repair / positive regulation of RNA splicing / regulation of DNA-templated transcription elongation / transcription elongation factor complex / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / response to gamma radiation / nucleotide-excision repair / transcription initiation at RNA polymerase II promoter / promoter-specific chromatin binding / P-body / Recognition of DNA damage by PCNA-containing replication complex / helicase activity 類似検索 - 分子機能 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.8 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Kokic, G. / Cramer, P. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 資金援助 |  ドイツ, 2件 ドイツ, 2件
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2021 タイトル: Structural basis of human transcription-DNA repair coupling. 著者: Goran Kokic / Felix R Wagner / Aleksandar Chernev / Henning Urlaub / Patrick Cramer / 要旨: Transcription-coupled DNA repair removes bulky DNA lesions from the genome and protects cells against ultraviolet (UV) irradiation. Transcription-coupled DNA repair begins when RNA polymerase II ...Transcription-coupled DNA repair removes bulky DNA lesions from the genome and protects cells against ultraviolet (UV) irradiation. Transcription-coupled DNA repair begins when RNA polymerase II (Pol II) stalls at a DNA lesion and recruits the Cockayne syndrome protein CSB, the E3 ubiquitin ligase, CRL4 and UV-stimulated scaffold protein A (UVSSA). Here we provide five high-resolution structures of Pol II transcription complexes containing human transcription-coupled DNA repair factors and the elongation factors PAF1 complex (PAF) and SPT6. Together with biochemical and published data, the structures provide a model for transcription-repair coupling. Stalling of Pol II at a DNA lesion triggers replacement of the elongation factor DSIF by CSB, which binds to PAF and moves upstream DNA to SPT6. The resulting elongation complex, EC, uses the CSA-stimulated translocase activity of CSB to pull on upstream DNA and push Pol II forward. If the lesion cannot be bypassed, CRL4 spans over the Pol II clamp and ubiquitylates the RPB1 residue K1268, enabling recruitment of TFIIH to UVSSA and DNA repair. Conformational changes in CRL4 lead to ubiquitylation of CSB and to release of transcription-coupled DNA repair factors before transcription may continue over repaired DNA. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7oo3.cif.gz | 2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7oo3.ent.gz | 1.6 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7oo3.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7oo3_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7oo3_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 7oo3_validation.xml.gz | 155 KB | 表示 | |
| CIF形式データ | 7oo3_validation.cif.gz | 251.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/oo/7oo3ftp://data.pdbj.org/pub/pdb/validation_reports/oo/7oo3 | HTTPS FTP |
-関連構造データ
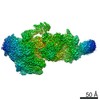








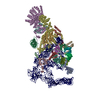




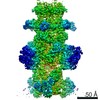
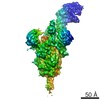

-リンク
 PDBj
PDBj










































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA-directed RNA polymerase II subunit ... , 6種, 6分子 ACEFGI
| #1: タンパク質 | 分子量: 217450.078 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #3: タンパク質 | 分子量: 31439.074 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質 | 分子量: 24644.318 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #6: タンパク質 | 分子量: 14477.001 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質 | 分子量: 19314.283 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 14541.221 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 6種, 6分子 BDKLcd
| #2: タンパク質 | 分子量: 134041.422 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #4: タンパク質 | 分子量: 16331.255 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 13310.284 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 7018.244 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #19: タンパク質 | 分子量: 80721.680 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: UVSSA, KIAA1530 / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q2YD98 Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q2YD98 |
| #20: タンパク質 | 分子量: 127369.719 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: DDB1, XAP1 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q16531 |
-DNA-directed RNA polymerases I, II, and III subunit ... , 2種, 2分子 HJ
| #8: タンパク質 | 分子量: 17162.273 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #10: タンパク質 | 分子量: 7655.123 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-DNA鎖 , 2種, 2分子 NT
| #13: DNA鎖 | 分子量: 14494.314 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #14: DNA鎖 | 分子量: 14269.129 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
-DNA excision repair protein ERCC- ... , 2種, 2分子 ab
| #16: タンパク質 | 分子量: 44107.160 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ERCC8, CKN1, CSA / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q13216 |
|---|---|
| #17: タンパク質 | 分子量: 168673.547 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ERCC6, CSB / 発現宿主: Trichoplusia ni (イラクサキンウワバ)参照: UniProt: Q03468, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 |
-RNA鎖 / タンパク質・ペプチド , 2種, 2分子 Px
| #15: RNA鎖 | 分子量: 3264.036 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #18: タンパク質・ペプチド | 分子量: 2996.685 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主: Trichoplusia ni (イラクサキンウワバ) |
-非ポリマー , 2種, 9分子 

| #21: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#22: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドのタイプ: Quantifoil R2/1 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 40.4 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 1 / 実像数: 10300 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.18.2_3874: / 分類: 精密化 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 2527839 | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 100000 詳細: Number of particles is different for every focused refined map, as detailed in the manuscript. 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 拘束条件 |
|
