 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6xmj | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Human 20S proteasome bound to an engineered 11S (PA26) activator | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | HYDROLASE / 11S-bound / 20S proteasome | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報proteasome activator complex / purine ribonucleoside triphosphate binding / regulation of endopeptidase activity / proteasome core complex / Regulation of ornithine decarboxylase (ODC) / Cross-presentation of soluble exogenous antigens (endosomes) / Somitogenesis / immune system process / myofibril / NF-kappaB binding ...proteasome activator complex / purine ribonucleoside triphosphate binding / regulation of endopeptidase activity / proteasome core complex / Regulation of ornithine decarboxylase (ODC) / Cross-presentation of soluble exogenous antigens (endosomes) / Somitogenesis / immune system process / myofibril / NF-kappaB binding / proteasome endopeptidase complex / proteasome core complex, beta-subunit complex / proteasome core complex, alpha-subunit complex / threonine-type endopeptidase activity / regulation of proteasomal protein catabolic process / negative regulation of inflammatory response to antigenic stimulus / : / sarcomere / proteasome complex / Regulation of activated PAK-2p34 by proteasome mediated degradation / ciliary basal body / proteolysis involved in protein catabolic process / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / Asymmetric localization of PCP proteins / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / Ubiquitin-dependent degradation of Cyclin D / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / Assembly of the pre-replicative complex / Vpu mediated degradation of CD4 / Degradation of DVL / Ubiquitin Mediated Degradation of Phosphorylated Cdc25A / Dectin-1 mediated noncanonical NF-kB signaling / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Hh mutants are degraded by ERAD / lipopolysaccharide binding / Degradation of AXIN / Degradation of GLI1 by the proteasome / Activation of NF-kappaB in B cells / Hedgehog ligand biogenesis / Defective CFTR causes cystic fibrosis / Negative regulation of NOTCH4 signaling / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / G2/M Checkpoints / Vif-mediated degradation of APOBEC3G / Autodegradation of the E3 ubiquitin ligase COP1 / Hedgehog 'on' state / Regulation of RUNX3 expression and activity / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / P-body / MAPK6/MAPK4 signaling / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / response to virus / Degradation of beta-catenin by the destruction complex / ABC-family proteins mediated transport / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / response to organic cyclic compound / CDK-mediated phosphorylation and removal of Cdc6 / CLEC7A (Dectin-1) signaling / SCF(Skp2)-mediated degradation of p27/p21 / Regulation of expression of SLITs and ROBOs / nuclear matrix / FCERI mediated NF-kB activation / Regulation of PTEN stability and activity / Interleukin-1 signaling / Orc1 removal from chromatin / Regulation of RAS by GAPs / Separation of Sister Chromatids / Regulation of RUNX2 expression and activity / The role of GTSE1 in G2/M progression after G2 checkpoint / UCH proteinases / KEAP1-NFE2L2 pathway / Antigen processing: Ubiquitination & Proteasome degradation / Downstream TCR signaling / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Neddylation / peptidase activity / positive regulation of NF-kappaB transcription factor activity / ER-Phagosome pathway / regulation of inflammatory response / postsynapse / proteasome-mediated ubiquitin-dependent protein catabolic process / secretory granule lumen / endopeptidase activity / response to oxidative stress / ficolin-1-rich granule lumen / nuclear body / ribosome / Ub-specific processing proteases / cadherin binding / intracellular membrane-bounded organelle / centrosome / ubiquitin protein ligase binding / synapse / Neutrophil degranulation / mitochondrion 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3 Å | |||||||||
 データ登録者 データ登録者 | de la Pena, A.H. / Opoku-Nsiah, K.A. / Williams, S.K. / Chopra, N. / Sali, A. / Gestwicki, J.E. / Lander, G.C. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2022 タイトル: The YΦ motif defines the structure-activity relationships of human 20S proteasome activators. 著者: Kwadwo A Opoku-Nsiah / Andres H de la Pena / Sarah K Williams / Nikita Chopra / Andrej Sali / Gabriel C Lander / Jason E Gestwicki / 要旨: The 20S proteasome (20S) facilitates turnover of most eukaryotic proteins. Substrate entry into the 20S first requires opening of gating loops through binding of HbYX motifs that are present at the C- ...The 20S proteasome (20S) facilitates turnover of most eukaryotic proteins. Substrate entry into the 20S first requires opening of gating loops through binding of HbYX motifs that are present at the C-termini of certain proteasome activators (PAs). The HbYX motif has been predominantly characterized in the archaeal 20S, whereas little is known about the sequence preferences of the human 20S (h20S). Here, we synthesize and screen ~120 HbYX-like peptides, revealing unexpected differences from the archaeal system and defining the h20S recognition sequence as the Y-F/Y (YФ) motif. To gain further insight, we create a functional chimera of the optimized sequence, NLSYYT, fused to the model activator, PA26. A cryo-EM structure of PA26-h20S is used to identify key interactions, including non-canonical contacts and gate-opening mechanisms. Finally, we demonstrate that the YФ sequence preferences are tuned by valency, allowing multivalent PAs to sample greater sequence space. These results expand the model for termini-mediated gating and provide a template for the design of h20S activators. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6xmj.cif.gz | 768.2 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6xmj.ent.gz | 626.4 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6xmj.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6xmj_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6xmj_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 6xmj_validation.xml.gz | 115.6 KB | 表示 | |
| CIF形式データ | 6xmj_validation.cif.gz | 179.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/xm/6xmjftp://data.pdbj.org/pub/pdb/validation_reports/xm/6xmj | HTTPS FTP |
-関連構造データ


-リンク
 PDBj
PDBj





- 集合体
集合体
| 登録構造単位 | 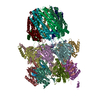
|
|---|---|
| 1 |
|
-要素
-Proteasome subunit alpha type- ... , 7種, 7分子 ABCDEFG
| #1: タンパク質 | 分子量: 27186.174 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA6, PROS27 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P60900, proteasome endopeptidase complex |
|---|---|
| #2: タンパク質 | 分子量: 25796.338 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA2, HC3, PSC3 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P25787, proteasome endopeptidase complex |
| #3: タンパク質 | 分子量: 28118.189 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA4, HC9, PSC9 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P25789, proteasome endopeptidase complex |
| #4: タンパク質 | 分子量: 27382.178 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA7, HSPC / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: O14818, proteasome endopeptidase complex |
| #5: タンパク質 | 分子量: 25569.957 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA5 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P28066, proteasome endopeptidase complex |
| #6: タンパク質 | 分子量: 26728.428 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA1, HC2, NU, PROS30, PSC2 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P25786, proteasome endopeptidase complex |
| #7: タンパク質 | 分子量: 27287.100 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMA3, HC8, PSC8 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P25788, proteasome endopeptidase complex |
-Proteasome subunit beta type- ... , 7種, 7分子 HIJKLMN
| #8: タンパク質 | 分子量: 21656.527 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB6, LMPY, Y / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P28072, proteasome endopeptidase complex |
|---|---|
| #9: タンパク質 | 分子量: 23745.256 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB7, Z / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q99436, proteasome endopeptidase complex |
| #10: タンパク質 | 分子量: 22841.701 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB3 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P49720, proteasome endopeptidase complex |
| #11: タンパク質 | 分子量: 22720.146 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB2 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P49721, proteasome endopeptidase complex |
| #12: タンパク質 | 分子量: 22199.072 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB5, LMPX, MB1, X / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P28074, proteasome endopeptidase complex |
| #13: タンパク質 | 分子量: 23578.986 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB1, PSC5 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P20618, proteasome endopeptidase complex |
| #14: タンパク質 | 分子量: 24138.453 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PSMB4, PROS26 / Cell (発現宿主): erythrocytes / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P28070, proteasome endopeptidase complex |
-タンパク質 , 1種, 7分子 OPQRSTU
| #15: タンパク質 | 分子量: 24959.373 Da / 分子数: 7 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主:  |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.6 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 20 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: HOMEMADE PLUNGER / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 277 K 詳細: Specimens were manually blotted with Whatman #1 filter paper. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 詳細: Images were acquired in nanoprobe mode. |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 29000 X / 最大 デフォーカス(公称値): -2500 nm / 最小 デフォーカス(公称値): -1000 nm / Calibrated defocus min: -1500 nm / 最大 デフォーカス(補正後): -3000 nm / Cs: 2.7 mm / C2レンズ絞り径: 70 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 6.25 sec. / 電子線照射量: 50 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 1 / 実像数: 11656 |
| 画像スキャン | サンプリングサイズ: 5 µm / 横: 3710 / 縦: 3838 / 動画フレーム数/画像: 25 / 利用したフレーム数/画像: 1-25 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 画像処理 | 詳細: Camera was operated in counting mode. | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | 詳細: CTF correction was performed by Relion during reconstruction. タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 579361 詳細: Particles were selected using the Relion template-based particle picker. | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 234960 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL |
