 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
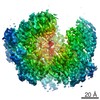
Yorodumi- PDB-6fe8: Cryo-EM structure of the core Centromere Binding Factor 3 complex -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6fe8 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of the core Centromere Binding Factor 3 complex | ||||||
 Components Components |
| ||||||
 Keywords Keywords | DNA BINDING PROTEIN / Centromere / CDEIII-binding / LRR domain | ||||||
| Function / homology |  Function and homology information Function and homology informationRAVE complex / Iron uptake and transport / : / CBF3 complex / regulation of transcription by galactose / cellular response to methylmercury / vacuolar proton-transporting V-type ATPase complex assembly / septin ring assembly / centromeric DNA binding / regulation of exit from mitosis ...RAVE complex / Iron uptake and transport / : / CBF3 complex / regulation of transcription by galactose / cellular response to methylmercury / vacuolar proton-transporting V-type ATPase complex assembly / septin ring assembly / centromeric DNA binding / regulation of exit from mitosis / kinetochore assembly / positive regulation of D-glucose transmembrane transport / vacuolar acidification / exit from mitosis / protein neddylation / mitochondrial fusion / mitotic intra-S DNA damage checkpoint signaling / silent mating-type cassette heterochromatin formation / DNA binding, bending / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / SCF ubiquitin ligase complex / mitotic spindle assembly checkpoint signaling / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Orc1 removal from chromatin / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Antigen processing: Ubiquitination & Proteasome degradation / DNA replication origin binding / cullin family protein binding / regulation of protein-containing complex assembly / subtelomeric heterochromatin formation / negative regulation of cytoplasmic translation / endomembrane system / regulation of mitotic cell cycle / G1/S transition of mitotic cell cycle / kinetochore / G2/M transition of mitotic cell cycle / mitotic cell cycle / protein-containing complex assembly / ubiquitin-dependent protein catabolic process / DNA-binding transcription factor activity, RNA polymerase II-specific / chromosome, telomeric region / protein ubiquitination / zinc ion binding / identical protein binding / nucleus / cytoplasm Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.1 Å | ||||||
 Authors Authors | Zhang, W.J. / Lukoynova, N. / Miah, S. / Vaughan, C.K. | ||||||
| Funding support |  United Kingdom, 1items United Kingdom, 1items
| ||||||
 Citation Citation | Journal: Cell Rep / Year: 2018 Title: Insights into Centromere DNA Bending Revealed by the Cryo-EM Structure of the Core Centromere Binding Factor 3 with Ndc10. Authors: Wenjuan Zhang / Natalya Lukoyanova / Shomon Miah / Jonathan Lucas / Cara K Vaughan / Abstract: The centromere binding factor 3 (CBF3) complex binds the third centromere DNA element in organisms with point centromeres, such as S. cerevisiae. It is an essential complex for assembly of the ...The centromere binding factor 3 (CBF3) complex binds the third centromere DNA element in organisms with point centromeres, such as S. cerevisiae. It is an essential complex for assembly of the kinetochore in these organisms, as it facilitates genetic centromere specification and allows association of all other kinetochore components. We determined high-resolution structures of the core complex of CBF3 alone and in association with a monomeric construct of Ndc10, using cryoelectron microscopy (cryo-EM). We identify the DNA-binding site of the complex and present a model in which CBF3 induces a tight bend in centromeric DNA, thus facilitating assembly of the centromeric nucleosome. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6fe8.cif.gz | 294.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6fe8.ent.gz | 228.1 KB | Display | PDB format |
| PDBx/mmJSON format | 6fe8.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/fe/6fe8ftp://data.pdbj.org/pub/pdb/validation_reports/fe/6fe8 | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 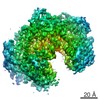 4241MC 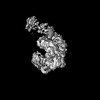 0051C  0052C 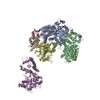 6gsaC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj







- Assembly
Assembly
| Deposited unit | 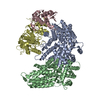
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 68454.125 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Details: Model is numbered according to Source: (gene. exp.) Gene: CEP3, CBF3, CBF3B, CSL1, YMR168C, YM8520.17C / Production host: #2: Protein | | Mass: 22558.451 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: SKP1, CBF3D, YDR328C, D9798.14 / Production host: #3: Protein | | Mass: 60899.961 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: CTF13, CBF3C, YMR094W, YM6543.01, YM9582.19 / Production host: Has protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: The core Centromere Binding Factor 3 complex / Type: COMPLEX Details: The core CBF3 complex, recombinantly expressed in Saccharomyces cerevisiae. It comprises a Cep3 homodimer, in which the binuclear zinc cluster domains are truncated, and full length Skp1 and Ctf13 components. Entity ID: all / Source: RECOMBINANT | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.22 MDa / Experimental value: NO | ||||||||||||||||||||
| Source (natural) | Organism: | ||||||||||||||||||||
| Source (recombinant) | Organism: | ||||||||||||||||||||
| Buffer solution | pH: 8 | ||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||
| Specimen | Conc.: 0.12 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: The sample is homogeneous and well-dispersed on grids. | ||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil R1.2/1.3 | ||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK I / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Microscopy | Model: FEI TITAN KRIOS | ||||||||||||
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM | ||||||||||||
| Electron lens | Mode: BRIGHT FIELD / Calibrated magnification: 47170 X / Cs: 2.7 mm | ||||||||||||
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER | ||||||||||||
| Image recording | Imaging-ID: 1 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 1
| ||||||||||||
| Image scans | Movie frames/image: 40 / Used frames/image: 1-40 |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.13_2992: / Classification: refinement | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
|