 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5of4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | The cryo-EM structure of human TFIIH | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | TRANSCRIPTION / transcription initiation / DNA repair / multiprotein complex / kinase / helicase | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報MMXD complex / core TFIIH complex portion of holo TFIIH complex / Cytosolic iron-sulfur cluster assembly / central nervous system myelin formation / positive regulation of mitotic recombination / hair cell differentiation / nucleotide-excision repair factor 3 complex / nucleotide-excision repair, preincision complex assembly / hair follicle maturation / CAK-ERCC2 complex ...MMXD complex / core TFIIH complex portion of holo TFIIH complex / Cytosolic iron-sulfur cluster assembly / central nervous system myelin formation / positive regulation of mitotic recombination / hair cell differentiation / nucleotide-excision repair factor 3 complex / nucleotide-excision repair, preincision complex assembly / hair follicle maturation / CAK-ERCC2 complex / embryonic cleavage / UV protection / transcription factor TFIIH core complex / transcription factor TFIIH holo complex / DNA 5'-3' helicase / G protein-coupled receptor internalization / transcription preinitiation complex / RNA Polymerase I Transcription Termination / transcription factor TFIID complex / RNA polymerase II general transcription initiation factor activity / erythrocyte maturation / regulation of mitotic cell cycle phase transition / hematopoietic stem cell proliferation / RNA Pol II CTD phosphorylation and interaction with CE during HIV infection / RNA Pol II CTD phosphorylation and interaction with CE / Formation of the Early Elongation Complex / Formation of the HIV-1 Early Elongation Complex / mRNA Capping / spinal cord development / bone mineralization / HIV Transcription Initiation / RNA Polymerase II HIV Promoter Escape / Transcription of the HIV genome / RNA Polymerase II Promoter Escape / RNA Polymerase II Transcription Pre-Initiation And Promoter Opening / RNA Polymerase II Transcription Initiation / RNA Polymerase II Transcription Initiation And Promoter Clearance / ATPase activator activity / 3'-5' DNA helicase activity / DNA 3'-5' helicase / DNA topological change / RNA Polymerase I Transcription Initiation / intrinsic apoptotic signaling pathway by p53 class mediator / hematopoietic stem cell differentiation / embryonic organ development / Tat-mediated elongation of the HIV-1 transcript / Formation of HIV-1 elongation complex containing HIV-1 Tat / transcription elongation by RNA polymerase I / response to UV / Formation of HIV elongation complex in the absence of HIV Tat / RNA Polymerase II Transcription Elongation / Formation of RNA Pol II elongation complex / extracellular matrix organization / transcription-coupled nucleotide-excision repair / RNA Polymerase II Pre-transcription Events / insulin-like growth factor receptor signaling pathway / DNA helicase activity / determination of adult lifespan / post-embryonic development / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / TP53 Regulates Transcription of DNA Repair Genes / nucleotide-excision repair / transcription initiation at RNA polymerase II promoter / promoter-specific chromatin binding / RNA Polymerase I Promoter Escape / transcription elongation by RNA polymerase II / chromosome segregation / cellular response to gamma radiation / NoRC negatively regulates rRNA expression / spindle / multicellular organism growth / Dual Incision in GG-NER / Transcription-Coupled Nucleotide Excision Repair (TC-NER) / Formation of TC-NER Pre-Incision Complex / Formation of Incision Complex in GG-NER / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / intracellular protein localization / 4 iron, 4 sulfur cluster binding / response to oxidative stress / double-stranded DNA binding / transcription by RNA polymerase II / 5'-3' DNA helicase activity / in utero embryonic development / damaged DNA binding / protein-macromolecule adaptor activity / response to hypoxia / nuclear speck / positive regulation of apoptotic process / DNA repair / apoptotic process / regulation of DNA-templated transcription / regulation of transcription by RNA polymerase II / nucleolus / ATP hydrolysis activity / DNA binding / zinc ion binding / nucleoplasm / ATP binding / metal ion binding 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.4 Å | |||||||||
 データ登録者 データ登録者 | Greber, B.J. / Nguyen, T.H.D. / Fang, J. / Afonine, P.V. / Adams, P.D. / Nogales, E. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Nature / 年: 2017 タイトル: The cryo-electron microscopy structure of human transcription factor IIH. 著者: Basil J Greber / Thi Hoang Duong Nguyen / Jie Fang / Pavel V Afonine / Paul D Adams / Eva Nogales / 要旨: Human transcription factor IIH (TFIIH) is part of the general transcriptional machinery required by RNA polymerase II for the initiation of eukaryotic gene transcription. Composed of ten subunits ...Human transcription factor IIH (TFIIH) is part of the general transcriptional machinery required by RNA polymerase II for the initiation of eukaryotic gene transcription. Composed of ten subunits that add up to a molecular mass of about 500 kDa, TFIIH is also essential for nucleotide excision repair. The seven-subunit TFIIH core complex formed by XPB, XPD, p62, p52, p44, p34, and p8 is competent for DNA repair, while the CDK-activating kinase subcomplex, which includes the kinase activity of CDK7 as well as the cyclin H and MAT1 subunits, is additionally required for transcription initiation. Mutations in the TFIIH subunits XPB, XPD, and p8 lead to severe premature ageing and cancer propensity in the genetic diseases xeroderma pigmentosum, Cockayne syndrome, and trichothiodystrophy, highlighting the importance of TFIIH for cellular physiology. Here we present the cryo-electron microscopy structure of human TFIIH at 4.4 Å resolution. The structure reveals the molecular architecture of the TFIIH core complex, the detailed structures of its constituent XPB and XPD ATPases, and how the core and kinase subcomplexes of TFIIH are connected. Additionally, our structure provides insight into the conformational dynamics of TFIIH and the regulation of its activity. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5of4.cif.gz | 459.1 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5of4.ent.gz | 356.8 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5of4.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/of/5of4ftp://data.pdbj.org/pub/pdb/validation_reports/of/5of4 | HTTPS FTP |
|---|
-関連構造データ

-リンク
 PDBj
PDBj













- 集合体
集合体
| 登録構造単位 | 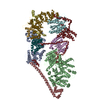
|
|---|---|
| 1 |
|
-要素
-TFIIH basal transcription factor complex helicase ... , 2種, 2分子 AB
| #1: タンパク質 | 分子量: 62426.047 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HeLa / 参照: UniProt: P19447, DNA helicase |
|---|---|
| #2: タンパク質 | 分子量: 87021.078 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HeLa / 参照: UniProt: P18074, DNA helicase |
-General transcription factor IIH subunit ... , 4種, 4分子 DEFG
| #3: タンパク質 | 分子量: 9875.401 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HeLa / 参照: UniProt: Q92759 |
|---|---|
| #4: タンパク質 | 分子量: 44481.996 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HeLa / 参照: UniProt: Q13888 |
| #5: タンパク質 | 分子量: 34416.008 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HeLa / 参照: UniProt: Q13889 |
| #6: タンパク質 | 分子量: 8060.362 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HeLa / 参照: UniProt: Q6ZYL4 |
-タンパク質 , 2種, 2分子 HZ
| #7: タンパク質 | 分子量: 10571.022 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Sequence register unassigned. / 由来: (天然) Homo sapiens (ヒト) |
|---|---|
| #8: タンパク質 | 分子量: 22996.232 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Sequence unassigned. / 由来: (天然) Homo sapiens (ヒト) |
-Unassigned secondary structure elements ... , 2種, 2分子 YX
| #9: タンパク質 | 分子量: 19762.281 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Sequence unassigned / 由来: (天然) Homo sapiens (ヒト) |
|---|---|
| #10: タンパク質 | 分子量: 6656.196 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) |
-非ポリマー , 1種, 1分子 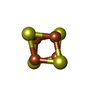
| #11: 化合物 | ChemComp-SF4 /  分子量: 351.640 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Fe4S4 分子量: 351.640 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Fe4S4 |
|---|
-詳細
| Has protein modification | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| ||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.8 | ||||||||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.0049 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: Natively purified complex at approx. 10 nM concentration | ||||||||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: C-flat-4/2 | ||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 278.15 K / 詳細: 3-4 minute incubation, 2 second blot |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 顕微鏡 | モデル: FEI TITAN |
|---|---|
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(補正後): 37879 X / 最大 デフォーカス(公称値): 4500 nm / 最小 デフォーカス(公称値): 2000 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: GATAN 626 SINGLE TILT LIQUID NITROGEN CRYO TRANSFER HOLDER |
| 撮影 | 平均露光時間: 8.7 sec. / 電子線照射量: 40 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 4 / 実像数: 8300 詳細: 8300 micrographs collected in four session with identical acquisition settings. Sessions lasted 4, 2, 4, and 4 days and yielded 1200, 1700, 2800, and 2600 micrographs. |
| 画像スキャン | サンプリングサイズ: 5 µm / 横: 3840 / 縦: 3710 / 動画フレーム数/画像: 30 / 利用したフレーム数/画像: 1-30 |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0158 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 画像処理 | 詳細: Micrographs were inspected for quality of Thon rings and ice contamination. Poor micrographs were rejected. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | 詳細: Correction in RELION based on values determined in CTFFIND4. タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1500000 詳細: Initial rounds of particle selection using DogPicker within APPION to generate templates for subsequent RELION autopicking. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 122900 / アルゴリズム: FOURIER SPACE / 詳細: RELION 3D auto-refinement (gold standard). / クラス平均像の数: 3 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL 詳細: Initial model assembled from high-resolution structures and homology models. Subsequently rebuilt in O and COOT, refined into the cryo-EM map using PHENIX and REFMAC, and fully validated. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 4.4→166.32 Å / Cor.coef. Fo:Fc: 0.939 / SU B: 54.849 / SU ML: 0.668 / ESU R: 1.679 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å / 溶媒モデル: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 193.578 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 17200 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|