 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4nh1: Crystal structure of a heterotetrameric CK2 holoenzyme complex ca... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4nh1 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal structure of a heterotetrameric CK2 holoenzyme complex carrying the Andante-mutation in CK2beta and consistent with proposed models of autoinhibition and trans-autophosphorylation | ||||||
 Components Components | (Casein kinase II subunit ...) x 2 | ||||||
 Keywords Keywords | TRANSFERASE / eukaryotic protein kinase fold / phosphotransferase (kinase) / phosphorylation | ||||||
| Function / homology |  Function and homology information Function and homology informationadiponectin-activated signaling pathway / positive regulation of activin receptor signaling pathway / endothelial tube morphogenesis / negative regulation of viral life cycle / protein kinase regulator activity / Phosphorylation and nuclear translocation of BMAL1 (ARNTL) and CLOCK / positive regulation of aggrephagy / regulation of chromosome separation / WNT mediated activation of DVL / Condensation of Prometaphase Chromosomes ...adiponectin-activated signaling pathway / positive regulation of activin receptor signaling pathway / endothelial tube morphogenesis / negative regulation of viral life cycle / protein kinase regulator activity / Phosphorylation and nuclear translocation of BMAL1 (ARNTL) and CLOCK / positive regulation of aggrephagy / regulation of chromosome separation / WNT mediated activation of DVL / Condensation of Prometaphase Chromosomes / protein kinase CK2 complex / symbiont-mediated disruption of host cell PML body / sperm glycocalyx / Phosphorylation and nuclear translocation of the CRY:PER:kinase complex / perinuclear theca / Regulation of CDH1 posttranslational processing and trafficking to plasma membrane / Receptor Mediated Mitophagy / Synthesis of PC / Sin3-type complex / negative regulation of signal transduction by p53 class mediator / RUNX1 interacts with co-factors whose precise effect on RUNX1 targets is not known / Maturation of hRSV A proteins / negative regulation of blood vessel endothelial cell migration / positive regulation of SMAD protein signal transduction / negative regulation of apoptotic signaling pathway / negative regulation of double-strand break repair via homologous recombination / positive regulation of Wnt signaling pathway / negative regulation of proteasomal ubiquitin-dependent protein catabolic process / Signal transduction by L1 / Hsp90 protein binding / PML body / fibrillar center / Wnt signaling pathway / Regulation of PTEN stability and activity / kinase activity / positive regulation of protein catabolic process / KEAP1-NFE2L2 pathway / rhythmic process / double-strand break repair / Cooperation of PDCL (PhLP1) and TRiC/CCT in G-protein beta folding / protein folding / positive regulation of cell growth / protein-containing complex assembly / sperm midpiece / secretory granule lumen / Regulation of TP53 Activity through Phosphorylation / ficolin-1-rich granule lumen / RNA polymerase II-specific DNA-binding transcription factor binding / protein-macromolecule adaptor activity / non-specific serine/threonine protein kinase / regulation of cell cycle / negative regulation of translation / protein stabilization / signaling receptor binding / negative regulation of cell population proliferation / protein domain specific binding / protein serine kinase activity / protein serine/threonine kinase activity / apoptotic process / positive regulation of cell population proliferation / chromatin binding / DNA damage response / Neutrophil degranulation / chromatin / signal transduction / extracellular exosome / extracellular region / nucleoplasm / ATP binding / metal ion binding / identical protein binding / nucleus / plasma membrane / cytoplasm / cytosol Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.3 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.3 Å | ||||||
 Authors Authors | Schnitzler, A. / Issinger, O.-G. / Niefind, K. | ||||||
 Citation Citation | Journal: J.Mol.Biol. / Year: 2014 Title: The Protein Kinase CK2(Andante) Holoenzyme Structure Supports Proposed Models of Autoregulation and Trans-Autophosphorylation Authors: Schnitzler, A. / Olsen, B.B. / Issinger, O.-G. / Niefind, K. #1: Journal: Mol.Cell.Biochem. / Year: 2005 Title: Biochemical characterization of the recombinant human Drosophila homologues Timekeeper and Andante involved in the Drosophila circadian oscillator. Authors: Rasmussen, T. / Skjth, I.H. / Jensen, H.H. / Niefind, K. / Boldyreff, B. / Issinger, O.G. #2: Journal: Embo J. / Year: 2001Title: Crystal structure of human protein kinase CK2: insights into basic properties of the CK2 holoenzyme. Authors: Niefind, K. / Guerra, B. / Ermakowa, I. / Issinger, O.G. #3: Journal: Mol.Cell.Biochem. / Year: 2005 Title: Primary and secondary interactions between CK2alpha and CK2beta lead to ring-like structures in the crystals of the CK2 holoenzyme. Authors: Niefind, K. / Issinger, O.G. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4nh1.cif.gz | 468.7 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4nh1.ent.gz | 391.5 KB | Display | PDB format |
| PDBx/mmJSON format | 4nh1.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/nh/4nh1ftp://data.pdbj.org/pub/pdb/validation_reports/nh/4nh1 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  1jwhS S: Starting model for refinement |
|---|---|
| Similar structure data |







-Links
 PDBj
PDBj







- Assembly
Assembly
| Deposited unit | 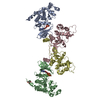
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
-Casein kinase II subunit ... , 2 types, 4 molecules ABCD
| #1: Protein | Mass: 40066.742 Da / Num. of mol.: 2 / Fragment: UNP residues 1-335 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CSNK2A1 / Plasmid: pT7-7 / Production host:  References: UniProt: P68400, non-specific serine/threonine protein kinase #2: Protein | Mass: 24951.375 Da / Num. of mol.: 2 / Mutation: Met166Ile Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CSNK2B / Plasmid: pT7-7 / Production host: |
|---|
-Non-polymers , 5 types, 29 molecules 
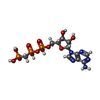
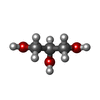


| #3: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg#4: Chemical |  Mass: 505.208 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C11H18N5O12P3 / Comment: AMP-PCP, energy-carrying molecule analogue*YM Mass: 505.208 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C11H18N5O12P3 / Comment: AMP-PCP, energy-carrying molecule analogue*YM#5: Chemical | ChemComp-GOL / |  Mass: 92.094 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C3H8O3 Mass: 92.094 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C3H8O3#6: Chemical |  Mass: 65.409 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Zn#7: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 20 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.85 Å3/Da / Density % sol: 56.8 % |
|---|
-Data collection
| Diffraction source | Source: SYNCHROTRON / Site: SLS  / Beamline: X06DA / Wavelength: 1 Å / Beamline: X06DA / Wavelength: 1 Å |
|---|---|
| Detector | Date: Jun 12, 2012 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 3.3→46.24 Å / Num. obs: 21878 / Redundancy: 2.4 % / Biso Wilson estimate: 106.58 Å2 / Rmerge(I) obs: 0.05 / Rsym value: 0.05 / Net I/σ(I): 13.85 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 1JWH Resolution: 3.3→46.237 Å / Occupancy max: 1 / Occupancy min: 0.29 / SU ML: 0.46 / σ(F): 1.35 / Phase error: 29.03 / Stereochemistry target values: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 308.81 Å2 / Biso mean: 130.8526 Å2 / Biso min: 27.82 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.3→46.237 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Refine-ID: X-RAY DIFFRACTION / Total num. of bins used: 8
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
