 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-7ebc: Crystal structure of Isocitrate lyase-1 from Saccaromyces cervisiae -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7ebc | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal structure of Isocitrate lyase-1 from Saccaromyces cervisiae | ||||||
 Components Components | Isocitrate lyase | ||||||
 Keywords Keywords | LYASE / Isocitrate lyase / glyoxylate cycle / ubiquitination | ||||||
| Function / homology |  Function and homology information Function and homology informationmethylisocitrate lyase / isocitrate lyase / isocitrate lyase activity / methylisocitrate lyase activity / glyoxylate cycle / vacuole / tricarboxylic acid cycle / extracellular region / metal ion binding / cytoplasm Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.3 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.3 Å | ||||||
 Authors Authors | Hiragi, K. / Nishio, K. / Moriyama, S. / Hamaguchi, T. / Mizoguchi, A. / Yonekura, K. / Tani, K. / Mizushima, T. | ||||||
 Citation Citation | Journal: J Struct Biol / Year: 2021 Title: Structural insights into the targeting specificity of ubiquitin ligase for S. cerevisiae isocitrate lyase but not C. albicans isocitrate lyase. Authors: Keito Hiragi / Kazuya Nishio / Shu Moriyama / Tasuku Hamaguchi / Akira Mizoguchi / Koji Yonekura / Kazutoshi Tani / Tsunehiro Mizushima /  Abstract: In Saccharomyces cerevisiae, the glyoxylate cycle is controlled through the posttranslational regulation of its component enzymes, such as isocitrate lyase (ICL), which catalyzes the first unique ...In Saccharomyces cerevisiae, the glyoxylate cycle is controlled through the posttranslational regulation of its component enzymes, such as isocitrate lyase (ICL), which catalyzes the first unique step of the cycle. The ICL of S.cerevisiae (ScIcl1) is tagged for proteasomal degradation through ubiquitination by a multisubunit ubiquitin ligase (the glucose-induced degradation-deficient (GID) complex), whereas that of the pathogenic yeast Candida albicans (CaIcl1) escapes this process. However, the reason for the ubiquitin targeting specificity of the GID complex for ScIcl1 and not for CaIcl1 is unclear. To gain some insight into this, in this study, the crystal structures of apo ScIcl1 and CaIcl1 in complex with formate and the cryogenic electron microscopy structure of apo CaIcl1 were determined at a resolution of 2.3, 2.7, and 2.6 Å, respectively. A comparison of the various structures suggests that the orientation of N-terminal helix α1 in S.cerevisiae is likely key to repositioning of ubiquitination sites and contributes to the distinction found in C. albicans ubiquitin evasion mechanism. This finding gives us a better understanding of the molecular mechanism of ubiquitin-dependent ScIcl1 degradation and could serve as a theoretical basis for the research and development of anti-C. albicans drugs based on the concept of CaIcl1 ubiquitination. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7ebc.cif.gz | 425.2 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7ebc.ent.gz | 345 KB | Display | PDB format |
| PDBx/mmJSON format | 7ebc.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/eb/7ebcftp://data.pdbj.org/pub/pdb/validation_reports/eb/7ebc | HTTPS FTP |
|---|
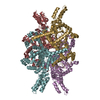
-Related structure data
| Related structure data | 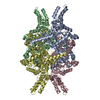 7ebeC 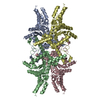 7ebfC  5e9gS S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |


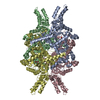

-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Unit cell |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS domain:
NCS domain segments: Component-ID: _ / Beg auth comp-ID: ASN / Beg label comp-ID: ASN / Refine code: _
NCS ensembles :
|
-Components
| #1: Protein | Mass: 63320.527 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Production host:  #2: Chemical | ChemComp-MG /   Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg#3: Chemical | ChemComp-PG4 / | 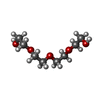  Mass: 194.226 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C8H18O5 / Comment: precipitant*YM Mass: 194.226 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C8H18O5 / Comment: precipitant*YM#4: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 575 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 575 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.75 Å3/Da / Density % sol: 55.24 % |
|---|---|
| Crystal grow | Temperature: 293.15 K / Method: vapor diffusion, sitting drop / Details: 40mM Na-HEPES pH7.0, 8%(w/v)PEG 3350 |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SPring-8 / Beamline: BL44XU / Wavelength: 0.9 Å |
| Detector | Type: DECTRIS EIGER X 16M / Detector: PIXEL / Date: Jun 7, 2019 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9 Å / Relative weight: 1 |
| Reflection | Resolution: 2.3→48.9 Å / Num. obs: 116065 / % possible obs: 99.7 % / Redundancy: 6.9 % / CC1/2: 0.997 / Net I/σ(I): 13.8 |
| Reflection shell | Resolution: 2.3→2.38 Å / Num. unique obs: 11263 / CC1/2: 0.788 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 5E9G Resolution: 2.3→48.8 Å / Cor.coef. Fo:Fc: 0.948 / Cor.coef. Fo:Fc free: 0.932 / SU B: 6.539 / SU ML: 0.153 / Cross valid method: THROUGHOUT / ESU R: 0.286 / ESU R Free: 0.204 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 37.349 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: 1 / Resolution: 2.3→48.8 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|