Netherlands Organisation for Scientific Research (NWO)
731.015.201
Netherlands
Netherlands Organisation for Scientific Research (NWO)
024.002.009
Netherlands
Citation
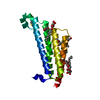
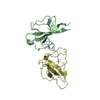
Journal: Life Sci Alliance / Year: 2020 Title: Implications for tetraspanin-enriched microdomain assembly based on structures of CD9 with EWI-F. Authors: Wout Oosterheert / Katerina T Xenaki / Viviana Neviani / Wouter Pos / Sofia Doulkeridou / Jip Manshande / Nicholas M Pearce / Loes Mj Kroon-Batenburg / Martin Lutz / Paul Mp van Bergen En ...Authors: Wout Oosterheert / Katerina T Xenaki / Viviana Neviani / Wouter Pos / Sofia Doulkeridou / Jip Manshande / Nicholas M Pearce / Loes Mj Kroon-Batenburg / Martin Lutz / Paul Mp van Bergen En Henegouwen / Piet Gros / Abstract: Tetraspanins are eukaryotic membrane proteins that contribute to a variety of signaling processes by organizing partner-receptor molecules in the plasma membrane. How tetraspanins bind and cluster ...Tetraspanins are eukaryotic membrane proteins that contribute to a variety of signaling processes by organizing partner-receptor molecules in the plasma membrane. How tetraspanins bind and cluster partner receptors into tetraspanin-enriched microdomains is unknown. Here, we present crystal structures of the large extracellular loop of CD9 bound to nanobodies 4C8 and 4E8 and, the cryo-EM structure of 4C8-bound CD9 in complex with its partner EWI-F. CD9-EWI-F displays a tetrameric arrangement with two central EWI-F molecules, dimerized through their ectodomains, and two CD9 molecules, one bound to each EWI-F transmembrane helix through CD9-helices h3 and h4. In the crystal structures, nanobodies 4C8 and 4E8 bind CD9 at loops C and D, which is in agreement with the 4C8 conformation in the CD9-EWI-F complex. The complex varies from nearly twofold symmetric (with the two CD9 copies nearly anti-parallel) to ca. 50° bent arrangements. This flexible arrangement of CD9-EWI-F with potential CD9 homo-dimerization at either end provides a "concatenation model" for forming short linear or circular assemblies, which may explain the occurrence of tetraspanin-enriched microdomains.
Mass: 10129.433 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CD9, MIC3, TSPAN29, GIG2 / Plasmid: pUPE 107.03 / Cell (production host): EMBRYONIC / Cell line (production host): HEK293EBNA / Organ (production host): KIDNEY / Production host: Homo sapiens (human) / References: UniProt: P21926
#2: Antibody
Nanobody4E8
Mass: 15546.274 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Lama glama (llama) / Production host: Escherichia coli BL21(DE3) (bacteria) / Variant (production host): Codon Plus
Mass: 18.015 Da / Num. of mol.: 134 / Source method: isolated from a natural source / Formula: H2O
-
Details
Has ligand of interest
N
Has protein modification
Y
-
Experimental details
-
Experiment
Experiment
Method: X-RAY DIFFRACTION / Number of used crystals: 1
-
Sample preparation
Crystal
Density Matthews: 1.91 Å3/Da / Density % sol: 35.67 % / Description: 3D-diamond like shape
Crystal grow
Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 8 Details: 0.2 M sodium acetate, 0.1 M Tris pH 8.0, 30% (w/v) PEG 4,000 cryoprotected with reservoir solution supplemented with 20% (v/v) ethylene glycol. Temp details: room temperature
-
Data collection
Diffraction
Mean temperature: 100 K / Ambient temp details: liquid nitrogen temperature / Serial crystal experiment: N
Method to determine structure: MOLECULAR REPLACEMENT Starting model: Structure of CD9EC2 bound to nanobody 4C8 Resolution: 1.33→44.72 Å / SU ML: 0.1369 / Cross valid method: FREE R-VALUE / σ(F): 1.35 / Phase error: 25.1549 / Stereochemistry target values: CDL v1.2 Details: For the CD9EC2 - 4E8 dataset, the autoprocessed and anisotropical-truncated (autoproc-staraniso) reflection data file provided by DLS was employed. The structure was solved by molecular ...Details: For the CD9EC2 - 4E8 dataset, the autoprocessed and anisotropical-truncated (autoproc-staraniso) reflection data file provided by DLS was employed. The structure was solved by molecular replacement using PHASER with the CD9EC2 - 4C8 structure as search model. The 4C8 residues were replaced with the corresponding 4E8 residues and the CDR regions of the nanobody were manually built in Coot. The structure was then iteratively refined using Refmac5 or Phenix, alternated with model improvement in COOT. The final refinement in Phenix yielded Rwork/Rfree = 15.1/19.0%
Rfactor
Num. reflection
% reflection
Rfree
0.1898
1570
4.9 %
Rwork
0.1512
30472
-
obs
0.1531
32042
69.74 %
Solvent computation
Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL
Displacement parameters
Biso mean: 27.7 Å2
Refinement step
Cycle: LAST / Resolution: 1.33→44.72 Å
Protein
Nucleic acid
Ligand
Solvent
Total
Num. atoms
1570
0
18
134
1722
Refine LS restraints
Refine-ID
Type
Dev ideal
Number
X-RAY DIFFRACTION
f_bond_d
0.0079
1775
X-RAY DIFFRACTION
f_angle_d
0.9406
2429
X-RAY DIFFRACTION
f_chiral_restr
0.0772
261
X-RAY DIFFRACTION
f_plane_restr
0.0057
319
X-RAY DIFFRACTION
f_dihedral_angle_d
23.8579
679
LS refinement shell
Resolution (Å)
Rfactor Rfree
Num. reflection Rfree
Rfactor Rwork
Num. reflection Rwork
Refine-ID
% reflection obs (%)
1.33-1.37
0.5263
20
0.2693
371
X-RAY DIFFRACTION
9.49
1.37-1.42
0.3523
45
0.2443
784
X-RAY DIFFRACTION
20.2
1.42-1.48
0.2764
78
0.211
1505
X-RAY DIFFRACTION
38.55
1.48-1.55
0.2478
125
0.181
2138
X-RAY DIFFRACTION
54.74
1.55-1.63
0.2464
150
0.1631
2820
X-RAY DIFFRACTION
72.09
1.63-1.73
0.2196
171
0.1594
3584
X-RAY DIFFRACTION
91.03
1.73-1.87
0.1857
203
0.1441
3957
X-RAY DIFFRACTION
99.93
1.87-2.05
0.1715
188
0.1322
3704
X-RAY DIFFRACTION
93.33
2.05-2.35
0.1893
189
0.1322
3436
X-RAY DIFFRACTION
86.56
2.35-2.96
0.1946
198
0.148
3956
X-RAY DIFFRACTION
97.67
2.96-44.72
0.1779
203
0.1578
4217
X-RAY DIFFRACTION
99.77
+
About Yorodumi
-
News
-
Feb 9, 2022. New format data for meta-information of EMDB entries
New format data for meta-information of EMDB entries
Version 3 of the EMDB header file is now the official format.
The previous official version 1.9 will be removed from the archive.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human)
 X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Netherlands, 2items
Netherlands, 2items  Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads











 PDBj
PDBj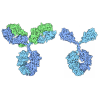

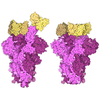
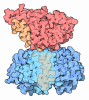


 Assembly
Assembly

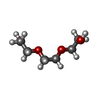

 Mass: 150.173 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H14O4
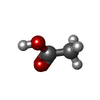
Mass: 150.173 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H14O4 Mass: 62.068 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C2H6O2
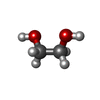
Mass: 62.068 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C2H6O2 Mass: 60.052 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C2H4O2
Mass: 60.052 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C2H4O2 Sample preparation
Sample preparation / Beamline: I04-1 / Wavelength: 0.9159 Å
/ Beamline: I04-1 / Wavelength: 0.9159 Å Processing
Processing