 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-6ftx: Structure of the chromatin remodelling enzyme Chd1 bound to a ubi... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6ftx | ||||||
|---|---|---|---|---|---|---|---|
| Title | Structure of the chromatin remodelling enzyme Chd1 bound to a ubiquitinylated nucleosome | ||||||
 Components Components |
| ||||||
 Keywords Keywords | MOTOR PROTEIN / Chromatin remodellers | ||||||
| Function / homology |  Function and homology information Function and homology informationregulation of transcriptional start site selection at RNA polymerase II promoter / nucleolar chromatin / regulation of chromatin organization / SLIK (SAGA-like) complex / negative regulation of DNA-templated DNA replication / DNA double-strand break processing / rDNA binding / nucleosome organization / nucleosome array spacer activity / histone H3K4me3 reader activity ...regulation of transcriptional start site selection at RNA polymerase II promoter / nucleolar chromatin / regulation of chromatin organization / SLIK (SAGA-like) complex / negative regulation of DNA-templated DNA replication / DNA double-strand break processing / rDNA binding / nucleosome organization / nucleosome array spacer activity / histone H3K4me3 reader activity / hypothalamus gonadotrophin-releasing hormone neuron development / female meiosis I / SAGA complex / ATP-dependent chromatin remodeler activity / seminiferous tubule development / positive regulation of protein monoubiquitination / fat pad development / mitochondrion transport along microtubule / sister chromatid cohesion / kinetochore assembly / female gonad development / termination of RNA polymerase II transcription / male meiosis I / termination of RNA polymerase I transcription / mitotic metaphase chromosome alignment / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / ATP-dependent activity, acting on DNA / energy homeostasis / nucleosomal DNA binding / neuron projection morphogenesis / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / regulation of neuron apoptotic process / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Prevention of phagosomal-lysosomal fusion / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / Negative regulation of FLT3 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / regulation of mitochondrial membrane potential / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Downregulation of ERBB4 signaling / APC-Cdc20 mediated degradation of Nek2A / Regulation of FZD by ubiquitination / p75NTR recruits signalling complexes / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / positive regulation of protein ubiquitination / NF-kB is activated and signals survival / regulation of proteasomal protein catabolic process / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / Pexophagy / Downregulation of ERBB2:ERBB3 signaling / NRIF signals cell death from the nucleus / Regulation of innate immune responses to cytosolic DNA / Regulation of PTEN localization / VLDLR internalisation and degradation / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by REV1 / TICAM1, RIP1-mediated IKK complex recruitment / Regulation of BACH1 activity / Translesion synthesis by POLK / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / InlB-mediated entry of Listeria monocytogenes into host cell / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Downregulation of TGF-beta receptor signaling / Translesion synthesis by POLI / Josephin domain DUBs / Gap-filling DNA repair synthesis and ligation in GG-NER / IKK complex recruitment mediated by RIP1 / PINK1-PRKN Mediated Mitophagy / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / TNFR1-induced NF-kappa-B signaling pathway / Regulation of activated PAK-2p34 by proteasome mediated degradation / TCF dependent signaling in response to WNT / Regulation of NF-kappa B signaling / activated TAK1 mediates p38 MAPK activation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / NOTCH3 Activation and Transmission of Signal to the Nucleus / Regulation of signaling by CBL / Negative regulators of DDX58/IFIH1 signaling / N-glycan trimming in the ER and Calnexin/Calreticulin cycle Similarity search - Function | ||||||
| Biological species |   Homo sapiens (human) Homo sapiens (human) synthetic construct (others) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.5 Å | ||||||
 Authors Authors | Sundaramoorthy, R. / Owen-hughes, T. / Norman, D.G. / Hughes, A. | ||||||
| Funding support |  United Kingdom, 1items United Kingdom, 1items
| ||||||
 Citation Citation | Journal: Elife / Year: 2018 Title: Structure of the chromatin remodelling enzyme Chd1 bound to a ubiquitinylated nucleosome. Authors: Ramasubramanian Sundaramoorthy / Amanda L Hughes / Hassane El-Mkami / David G Norman / Helder Ferreira / Tom Owen-Hughes / Abstract: ATP-dependent chromatin remodelling proteins represent a diverse family of proteins that share ATPase domains that are adapted to regulate protein-DNA interactions. Here, we present structures of the ...ATP-dependent chromatin remodelling proteins represent a diverse family of proteins that share ATPase domains that are adapted to regulate protein-DNA interactions. Here, we present structures of the Chd1 protein engaged with nucleosomes in the presence of the transition state mimic ADP-beryllium fluoride. The path of DNA strands through the ATPase domains indicates the presence of contacts conserved with single strand translocases and additional contacts with both strands that are unique to Snf2 related proteins. The structure provides connectivity between rearrangement of ATPase lobes to a closed, nucleotide bound state and the sensing of linker DNA. Two turns of linker DNA are prised off the surface of the histone octamer as a result of Chd1 binding, and both the histone H3 tail and ubiquitin conjugated to lysine 120 are re-orientated towards the unravelled DNA. This indicates how changes to nucleosome structure can alter the way in which histone epitopes are presented. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6ftx.cif.gz | 545.4 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6ftx.ent.gz | 420.5 KB | Display | PDB format |
| PDBx/mmJSON format | 6ftx.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ft/6ftxftp://data.pdbj.org/pub/pdb/validation_reports/ft/6ftx | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4318MC  4336C  6g0lC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj


















































- Assembly
Assembly
| Deposited unit | 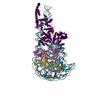
|
|---|---|
| 1 |
|
-Components
-Protein , 7 types, 11 molecules ABFCGDHENOW
| #1: Protein | Mass: 11431.358 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 11394.426 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #3: Protein | Mass: 14093.436 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #4: Protein | Mass: 13939.228 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: LOC108648866 / Production host: #5: Protein | | Mass: 12329.330 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #8: Protein | Mass: 8576.831 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: UBB / Production host: #9: Protein | | Mass: 102152.859 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: CHD1, SCKG_4184 / Production host: |
-DNA chain , 2 types, 2 molecules IJ
| #6: DNA chain | Mass: 48792.086 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
|---|---|
| #7: DNA chain | Mass: 49678.613 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
-Non-polymers , 2 types, 2 molecules 

| #10: Chemical | ChemComp-BEF /  Mass: 66.007 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: BeF3 Mass: 66.007 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: BeF3 |
|---|---|
| #11: Chemical | ChemComp-ADP /  Mass: 427.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM |
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.450 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | ||||||||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 1 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: Sample was gel filtration purified and it is monodisperse | ||||||||||||||||||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: C-flat-1.2/1.3 4C | ||||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK III / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 35714 X / Calibrated magnification: 35714 X / Nominal defocus max: 4000 nm / Nominal defocus min: 1500 nm / Calibrated defocus min: 1500 nm / Calibrated defocus max: 4000 nm / Cs: 2.7 mm / C2 aperture diameter: 70 µm / Alignment procedure: BASIC |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Temperature (max): 170 K / Temperature (min): 170 K |
| Image recording | Average exposure time: 0.32 sec. / Electron dose: 1.25 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 QUANTUM (4k x 4k) / Num. of grids imaged: 1 / Num. of real images: 1300 |
| EM imaging optics | Energyfilter name: GIF Quantum LS |
| Image scans | Sampling size: 5 µm / Width: 3870 / Height: 3870 / Movie frames/image: 32 / Used frames/image: 5-28 |
- Processing
Processing
| Software | Name: REFMAC / Version: 5.8.0230 / Classification: refinement | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 860000 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.5 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 135000 / Algorithm: FOURIER SPACE / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | B value: 204 / Protocol: FLEXIBLE FIT / Space: RECIPROCAL / Target criteria: Cross-correlation coefficient | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Cor.coef. Fo:Fc: 0.304 / Highest resolution: 4.5 Å / % reflection obs: 100 % / SU B: 200.492 / SU ML: 0.762 / ESU R: 0.624 Stereochemistry target values: MAXIMUM LIKELIHOOD WITH PHASES Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Solvent model: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 266.837 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: 1 / Total: 21021 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|