 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5i0i: Crystal structure of myosin X motor domain with 2IQ motifs in pre... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5i0i | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of myosin X motor domain with 2IQ motifs in pre-powerstroke state | |||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||
 Keywords Keywords | MOTOR PROTEIN / myosin / motor domain / molecular motor / pre-powerstrocke state / motility | |||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationplus-end directed microfilament motor activity / Netrin-1 signaling / positive regulation of cell-cell adhesion / cytoskeleton-dependent intracellular transport / : / : / : / : / filopodium tip / positive regulation of protein autophosphorylation ...plus-end directed microfilament motor activity / Netrin-1 signaling / positive regulation of cell-cell adhesion / cytoskeleton-dependent intracellular transport / : / : / : / : / filopodium tip / positive regulation of protein autophosphorylation / : / regulation of filopodium assembly / negative regulation of peptidyl-threonine phosphorylation / : / type 3 metabotropic glutamate receptor binding / filopodium membrane / myosin complex / positive regulation of DNA binding / CaM pathway / Cam-PDE 1 activation / positive regulation of peptidyl-threonine phosphorylation / Sodium/Calcium exchangers / Calmodulin induced events / Reduction of cytosolic Ca++ levels / Activation of Ca-permeable Kainate Receptor / CREB1 phosphorylation through the activation of CaMKII/CaMKK/CaMKIV cascasde / Loss of phosphorylation of MECP2 at T308 / CREB1 phosphorylation through the activation of Adenylate Cyclase / negative regulation of high voltage-gated calcium channel activity / PKA activation / CaMK IV-mediated phosphorylation of CREB / spectrin binding / Glycogen breakdown (glycogenolysis) / microfilament motor activity / response to corticosterone / negative regulation of ryanodine-sensitive calcium-release channel activity / Activation of RAC1 downstream of NMDARs / organelle localization by membrane tethering / CLEC7A (Dectin-1) induces NFAT activation / : / autophagosome membrane docking / regulation of synaptic vesicle exocytosis / negative regulation of calcium ion export across plasma membrane / regulation of cardiac muscle cell action potential / presynaptic endocytosis / Synthesis of IP3 and IP4 in the cytosol / positive regulation of protein serine/threonine kinase activity / Phase 0 - rapid depolarisation / Negative regulation of NMDA receptor-mediated neuronal transmission / phosphatidylinositol-3,4,5-trisphosphate binding / Unblocking of NMDA receptors, glutamate binding and activation / calcineurin-mediated signaling / RHO GTPases activate PAKs / nitric-oxide synthase binding / regulation of cell communication by electrical coupling involved in cardiac conduction / Ion transport by P-type ATPases / adenylate cyclase binding / Uptake and function of anthrax toxins / protein phosphatase activator activity / regulation of ryanodine-sensitive calcium-release channel activity / Long-term potentiation / Calcineurin activates NFAT / Regulation of MECP2 expression and activity / DARPP-32 events / Smooth Muscle Contraction / regulation of synaptic vesicle endocytosis / detection of calcium ion / catalytic complex / regulation of cardiac muscle contraction / cellular response to interferon-beta / RHO GTPases activate IQGAPs / positive regulation of nitric-oxide synthase activity / phosphatidylinositol 3-kinase binding / activation of adenylate cyclase activity / calcium channel inhibitor activity / presynaptic cytosol / Activation of AMPK downstream of NMDARs / regulation of release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / enzyme regulator activity / ruffle / eNOS activation / Ion homeostasis / Tetrahydrobiopterin (BH4) synthesis, recycling, salvage and regulation / regulation of calcium-mediated signaling / Protein methylation / titin binding / regulation of cardiac muscle contraction by regulation of the release of sequestered calcium ion / voltage-gated potassium channel complex / FCERI mediated Ca+2 mobilization / calcium channel complex / substantia nigra development / regulation of heart rate / FCGR3A-mediated IL10 synthesis / calyx of Held / Ras activation upon Ca2+ influx through NMDA receptor / Antigen activates B Cell Receptor (BCR) leading to generation of second messengers / response to amphetamine / nitric-oxide synthase regulator activity / adenylate cyclase activator activity / protein serine/threonine kinase activator activity Similarity search - Function | |||||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||||||||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.15 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.15 Å | |||||||||||||||||||||
 Authors Authors | Isabet, T. / Sweeney, H.L. / Houdusse, A. | |||||||||||||||||||||
| Funding support |  France, France,  United States, 6items United States, 6items
| |||||||||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2016 Title: The myosin X motor is optimized for movement on actin bundles. Authors: Virginie Ropars / Zhaohui Yang / Tatiana Isabet / Florian Blanc / Kaifeng Zhou / Tianming Lin / Xiaoyan Liu / Pascale Hissier / Frédéric Samazan / Béatrice Amigues / Eric D Yang / Hyokeun ...Authors: Virginie Ropars / Zhaohui Yang / Tatiana Isabet / Florian Blanc / Kaifeng Zhou / Tianming Lin / Xiaoyan Liu / Pascale Hissier / Frédéric Samazan / Béatrice Amigues / Eric D Yang / Hyokeun Park / Olena Pylypenko / Marco Cecchini / Charles V Sindelar / H Lee Sweeney / Anne Houdusse /  Abstract: Myosin X has features not found in other myosins. Its structure must underlie its unique ability to generate filopodia, which are essential for neuritogenesis, wound healing, cancer metastasis and ...Myosin X has features not found in other myosins. Its structure must underlie its unique ability to generate filopodia, which are essential for neuritogenesis, wound healing, cancer metastasis and some pathogenic infections. By determining high-resolution structures of key components of this motor, and characterizing the in vitro behaviour of the native dimer, we identify the features that explain the myosin X dimer behaviour. Single-molecule studies demonstrate that a native myosin X dimer moves on actin bundles with higher velocities and takes larger steps than on single actin filaments. The largest steps on actin bundles are larger than previously reported for artificially dimerized myosin X constructs or any other myosin. Our model and kinetic data explain why these large steps and high velocities can only occur on bundled filaments. Thus, myosin X functions as an antiparallel dimer in cells with a unique geometry optimized for movement on actin bundles. | |||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5i0i.cif.gz | 794.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5i0i.ent.gz | 653.1 KB | Display | PDB format |
| PDBx/mmJSON format | 5i0i.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/i0/5i0iftp://data.pdbj.org/pub/pdb/validation_reports/i0/5i0i | HTTPS FTP |
|---|
-Related structure data










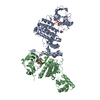
-Links
 PDBj
PDBj























- Assembly
Assembly
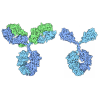
| Deposited unit | 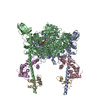
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| Unit cell |
|
-Components
-Protein , 4 types, 5 molecules ABCEI
| #1: Protein | Mass: 91550.148 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: MYO10, KIAA0799 / Production host:  unidentified baculovirus / References: UniProt: Q9HD67 unidentified baculovirus / References: UniProt: Q9HD67#2: Protein | | Mass: 16450.014 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: CALM1, CALM, CAM, CAM1, CALM2, CAM2, CAMB, CALM3, CALML2, CAM3, CAMC, CAMIII Production host: unidentified baculovirus / References: UniProt: P62158, UniProt: P0DP23*PLUS#3: Protein | | Mass: 16373.917 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: CALM1, CALM, CAM, CAM1, CALM2, CAM2, CAMB, CALM3, CALML2, CAM3, CAMC, CAMIII Production host: unidentified baculovirus / References: UniProt: P62158, UniProt: P0DP23*PLUS#5: Protein | | Mass: 7408.098 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: CALM1, CALM, CAM, CAM1, CALM2, CAM2, CAMB, CALM3, CALML2, CAM3, CAMC, CAMIII Production host: unidentified baculovirus / References: UniProt: P62158, UniProt: P0DP23*PLUS |
|---|
-Protein/peptide , 1 types, 1 molecules G
| #4: Protein/peptide | Mass: 4977.496 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: CALM1, CALM, CAM, CAM1, CALM2, CAM2, CAMB, CALM3, CALML2, CAM3, CAMC, CAMIII Production host: unidentified baculovirus / References: UniProt: P62158, UniProt: P0DP23*PLUS |
|---|
-Non-polymers , 6 types, 32 molecules 

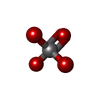

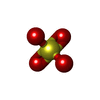

| #6: Chemical |  Mass: 209.263 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C7H15NO4S / Comment: pH buffer*YM Mass: 209.263 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C7H15NO4S / Comment: pH buffer*YM#7: Chemical |  Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg#8: Chemical |  Mass: 114.939 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: VO4 Mass: 114.939 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: VO4#9: Chemical |  Mass: 427.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM#10: Chemical | ChemComp-SO4 /  Mass: 96.063 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Formula: SO4 Mass: 96.063 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Formula: SO4#11: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 16 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.84 Å3/Da / Density % sol: 68 % |
|---|---|
| Crystal grow | Temperature: 277 K / Method: vapor diffusion, hanging drop / pH: 7 Details: 7.5% PEG 10000, 100mM MOPS pH 7.0, 1mM TCEP and 50mM Magnesium acetate |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SOLEIL / Beamline: PROXIMA 1 / Wavelength: 0.9801 Å |
| Detector | Type: DECTRIS PILATUS 6M / Detector: PIXEL / Date: Apr 9, 2013 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9801 Å / Relative weight: 1 |
| Reflection | Resolution: 3.15→50 Å / Num. obs: 61853 / % possible obs: 99.9 % / Redundancy: 16.9 % / Biso Wilson estimate: 118.26 Å2 / Net I/σ(I): 14.48 |
| Reflection shell | Resolution: 3.15→3.23 Å / Redundancy: 16.8 % / Mean I/σ(I) obs: 1.15 / % possible all: 99.7 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 3.15→49.53 Å / Cor.coef. Fo:Fc: 0.9356 / Cor.coef. Fo:Fc free: 0.9334 / SU R Cruickshank DPI: 3.519 / Cross valid method: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 1.802 / SU Rfree Blow DPI: 0.333 / SU Rfree Cruickshank DPI: 0.342
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 126.34 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.383 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.15→49.53 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 3.15→3.23 Å / Total num. of bins used: 20
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
