 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5aqg | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Fragment-based screening of HSP70 sheds light on the functional role of ATP-binding site residues | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | CHAPERONE / HEAT SHOCK PROTEIN / HSP70 / HSP72 / HSC70 / ATPASE / BAG1 / FRAGMENT | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報lumenal side of lysosomal membrane / regulation of protein import / negative regulation of supramolecular fiber organization / chaperone-mediated autophagy translocation complex disassembly / clathrin-sculpted gamma-aminobutyric acid transport vesicle membrane / Lipophagy / Respiratory syncytial virus genome transcription / GABA synthesis, release, reuptake and degradation / protein targeting to lysosome involved in chaperone-mediated autophagy / protein carrier chaperone ...lumenal side of lysosomal membrane / regulation of protein import / negative regulation of supramolecular fiber organization / chaperone-mediated autophagy translocation complex disassembly / clathrin-sculpted gamma-aminobutyric acid transport vesicle membrane / Lipophagy / Respiratory syncytial virus genome transcription / GABA synthesis, release, reuptake and degradation / protein targeting to lysosome involved in chaperone-mediated autophagy / protein carrier chaperone / adenyl-nucleotide exchange factor activity / positive regulation of smooth muscle cell apoptotic process / clathrin coat disassembly / C3HC4-type RING finger domain binding / CHL1 interactions / negative regulation of NLRP3 inflammasome complex assembly / regulation of protein complex stability / ATP-dependent protein disaggregase activity / membrane organization / protein folding chaperone complex / Lysosome Vesicle Biogenesis / cellular response to steroid hormone stimulus / Golgi Associated Vesicle Biogenesis / chaperone-mediated autophagy / non-chaperonin molecular chaperone ATPase / : / Prp19 complex / HSF1-dependent transactivation / Regulation of HSF1-mediated heat shock response / response to unfolded protein / regulation of protein-containing complex assembly / Attenuation phase / ATP metabolic process / Protein methylation / heat shock protein binding / protein folding chaperone / mRNA Splicing - Major Pathway / lysosomal lumen / AUF1 (hnRNP D0) binds and destabilizes mRNA / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / cellular response to starvation / Late endosomal microautophagy / spliceosomal complex / ATP-dependent protein folding chaperone / G protein-coupled receptor binding / mRNA splicing, via spliceosome / PKR-mediated signaling / Chaperone Mediated Autophagy / regulation of protein stability / MHC class II protein complex binding / unfolded protein binding / Clathrin-mediated endocytosis / melanosome / protein folding / protein-folding chaperone binding / protein refolding / blood microparticle / secretory granule lumen / Interleukin-4 and Interleukin-13 signaling / protein-macromolecule adaptor activity / ficolin-1-rich granule lumen / cell surface receptor signaling pathway / protein stabilization / positive regulation of cell migration / cadherin binding / receptor ligand activity / ribonucleoprotein complex / lysosomal membrane / focal adhesion / negative regulation of DNA-templated transcription / ubiquitin protein ligase binding / apoptotic process / Neutrophil degranulation / negative regulation of apoptotic process / nucleolus / enzyme binding / ATP hydrolysis activity / extracellular space / RNA binding / extracellular exosome / extracellular region / nucleoplasm / ATP binding / nucleus / membrane / plasma membrane / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  HOMO SAPIENS (ヒト) HOMO SAPIENS (ヒト) | ||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 2.24 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 2.24 Å | ||||||
 データ登録者 データ登録者 | Jones, A.M. / Westwood, I.M. / Osborne, J.D. / Matthews, T.P. / Cheeseman, M.D. / Rowlands, M.G. / Jeganathan, F. / Burke, R. / Lee, D. / Kadi, N. ...Jones, A.M. / Westwood, I.M. / Osborne, J.D. / Matthews, T.P. / Cheeseman, M.D. / Rowlands, M.G. / Jeganathan, F. / Burke, R. / Lee, D. / Kadi, N. / Liu, M. / Richards, M. / McAndrew, C. / Yahya, N. / Dobson, S.E. / Jones, K. / Workman, P. / Collins, I. / van Montfort, R.L.M. | ||||||
 引用 引用 | ジャーナル: Sci Rep / 年: 2016 タイトル: A fragment-based approach applied to a highly flexible target: Insights and challenges towards the inhibition of HSP70 isoforms. 著者: Jones, A.M. / Westwood, I.M. / Osborne, J.D. / Matthews, T.P. / Cheeseman, M.D. / Rowlands, M.G. / Jeganathan, F. / Burke, R. / Lee, D. / Kadi, N. / Liu, M. / Richards, M. / McAndrew, C. / ...著者: Jones, A.M. / Westwood, I.M. / Osborne, J.D. / Matthews, T.P. / Cheeseman, M.D. / Rowlands, M.G. / Jeganathan, F. / Burke, R. / Lee, D. / Kadi, N. / Liu, M. / Richards, M. / McAndrew, C. / Yahya, N. / Dobson, S.E. / Jones, K. / Workman, P. / Collins, I. / van Montfort, R.L. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5aqg.cif.gz | 583 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5aqg.ent.gz | 480.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5aqg.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5aqg_validation.pdf.gz | 1.3 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5aqg_full_validation.pdf.gz | 1.3 MB | 表示 | |
| XML形式データ | 5aqg_validation.xml.gz | 55.6 KB | 表示 | |
| CIF形式データ | 5aqg_validation.cif.gz | 78.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/aq/5aqgftp://data.pdbj.org/pub/pdb/validation_reports/aq/5aqg | HTTPS FTP |
-関連構造データ
| 関連構造データ |  5aqfC  5aqhC  5aqiC  5aqjC  5aqkC  5aqlC  5aqmC  5aqnC  5aqoC  5aqpC  5aqqC  5aqrC  5aqsC  5aqtC  5aquC  5aqvC  5aqwC  5aqxC  5aqyC  1hx1S C: 同じ文献を引用 ( S: 精密化の開始モデル |
|---|---|
| 類似構造データ |



-リンク
 PDBj
PDBj















- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 , 2種, 6分子 ACEBDF
| #1: タンパク質 | 分子量: 42406.980 Da / 分子数: 3 / 断片: NUCLEOTIDE BINDING DOMAIN, RESIDUES 1-381 / 由来タイプ: 組換発現 / 由来: (組換発現) HOMO SAPIENS (ヒト) / 発現宿主:  #2: タンパク質 | 分子量: 13511.571 Da / 分子数: 3 / 断片: RESIDUES 222-334 / 由来タイプ: 組換発現 / 由来: (組換発現) HOMO SAPIENS (ヒト) / 発現宿主: |
|---|
-非ポリマー , 5種, 484分子 
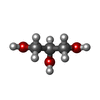
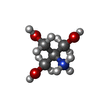


| #3: 化合物 |  分子量: 320.304 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C13H16N6O4 分子量: 320.304 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C13H16N6O4#4: 化合物 | ChemComp-GOL /  分子量: 92.094 Da / 分子数: 12 / 由来タイプ: 合成 / 式: C3H8O3 分子量: 92.094 Da / 分子数: 12 / 由来タイプ: 合成 / 式: C3H8O3#5: 化合物 |  分子量: 122.143 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C4H12NO3 / コメント: pH緩衝剤*YM 分子量: 122.143 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C4H12NO3 / コメント: pH緩衝剤*YM#6: 化合物 |  分子量: 35.453 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Cl 分子量: 35.453 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Cl#7: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 465 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.46 Å3/Da / 溶媒含有率: 49.92 % / 解説: NONE |
|---|---|
| 結晶化 | pH: 8.5 詳細: 16-26% (W/V) PEG3350, 0.1 M K-NA TARTRATE, 0.1 M TRIS.HCL PH 8.5 AND 25% (V/V) GLYCEROL |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: Diamond  / ビームライン: I24 / 波長: 0.9686 / ビームライン: I24 / 波長: 0.9686 |
| 検出器 | タイプ: DECTRIS PILATUS 6M / 検出器: PIXEL / 日付: 2012年7月29日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.9686 Å / 相対比: 1 |
| 反射 | 解像度: 2.24→47.59 Å / Num. obs: 78546 / % possible obs: 98.4 % / Observed criterion σ(I): 0 / 冗長度: 3.3 % / Biso Wilson estimate: 34.87 Å2 / Rmerge(I) obs: 0.11 / Net I/σ(I): 5.1 |
| 反射 シェル | 解像度: 2.24→2.28 Å / 冗長度: 2.1 % / Rmerge(I) obs: 0.96 / Mean I/σ(I) obs: 0.4 / % possible all: 84.3 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: PDB ENTRY 1HX1 解像度: 2.24→43.4 Å / Cor.coef. Fo:Fc: 0.934 / Cor.coef. Fo:Fc free: 0.92 / SU R Cruickshank DPI: 0.266 / 交差検証法: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.267 / SU Rfree Blow DPI: 0.192 / SU Rfree Cruickshank DPI: 0.194 詳細: IDEAL-DIST CONTACT TERM CONTACT SETUP. ALL ATOMS HAVE CCP4 ATOM TYPE FROM LIBRARY
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 48.14 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.306 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.24→43.4 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | 解像度: 2.24→2.3 Å / Total num. of bins used: 20
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
