 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4wrn | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
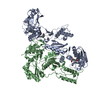
| タイトル | Crystal structure of the polymerization region of human uromodulin/Tamm-Horsfall protein | ||||||||||||||||||||||||
 要素 要素 | Maltose-binding periplasmic protein,Uromodulin | ||||||||||||||||||||||||
 キーワード キーワード | STRUCTURAL PROTEIN / ZP DOMAIN / EGF DOMAIN / EXTRACELLULAR MATRIX / GLYCOPROTEIN | ||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報citric acid secretion / metanephric thick ascending limb development / metanephric distal convoluted tubule development / connective tissue replacement / protein transport into plasma membrane raft / Asparagine N-linked glycosylation / organ or tissue specific immune response / collecting duct development / metanephric ascending thin limb development / micturition ...citric acid secretion / metanephric thick ascending limb development / metanephric distal convoluted tubule development / connective tissue replacement / protein transport into plasma membrane raft / Asparagine N-linked glycosylation / organ or tissue specific immune response / collecting duct development / metanephric ascending thin limb development / micturition / urea transmembrane transport / protein localization to vacuole / regulation of protein transport / juxtaglomerular apparatus development / antibacterial innate immune response / intracellular chloride ion homeostasis / renal urate salt excretion / urate transport / glomerular filtration / renal sodium ion absorption / neutrophil migration / response to water deprivation / intracellular phosphate ion homeostasis / potassium ion homeostasis / regulation of urine volume / intracellular sodium ion homeostasis / endoplasmic reticulum organization / IgG binding / heterophilic cell-cell adhesion / extrinsic component of membrane / ciliary membrane / leukocyte cell-cell adhesion / detection of maltose stimulus / maltose transport complex / carbohydrate transport / carbohydrate transmembrane transporter activity / maltose binding / cellular response to unfolded protein / maltose transport / maltodextrin transmembrane transport / cellular defense response / multicellular organismal response to stress / renal water homeostasis / side of membrane / ATP-binding cassette (ABC) transporter complex, substrate-binding subunit-containing / : / ERAD pathway / ATP-binding cassette (ABC) transporter complex / RNA splicing / tumor necrosis factor-mediated signaling pathway / cell chemotaxis / apoptotic signaling pathway / lipid metabolic process / autophagy / Golgi lumen / regulation of blood pressure / intracellular calcium ion homeostasis / spindle pole / outer membrane-bounded periplasmic space / response to lipopolysaccharide / defense response to Gram-negative bacterium / basolateral plasma membrane / periplasmic space / cilium / apical plasma membrane / response to xenobiotic stimulus / inflammatory response / negative regulation of cell population proliferation / calcium ion binding / DNA damage response / cell surface / endoplasmic reticulum / extracellular space / extracellular exosome / membrane 類似検索 - 分子機能 | ||||||||||||||||||||||||
| 生物種 |   Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||||||||||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 3.2 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 3.2 Å | ||||||||||||||||||||||||
 データ登録者 データ登録者 | Bokhove, M. / De Sanctis, D. / Jovine, L. | ||||||||||||||||||||||||
| 資金援助 |  スウェーデン, 7件 スウェーデン, 7件
| ||||||||||||||||||||||||
 引用 引用 | ジャーナル: Proc Natl Acad Sci U S A / 年: 2016 タイトル: A structured interdomain linker directs self-polymerization of human uromodulin. 著者: Marcel Bokhove / Kaoru Nishimura / Martina Brunati / Ling Han / Daniele de Sanctis / Luca Rampoldi / Luca Jovine /   要旨: Uromodulin (UMOD)/Tamm-Horsfall protein, the most abundant human urinary protein, plays a key role in chronic kidney diseases and is a promising therapeutic target for hypertension. Via its bipartite ...Uromodulin (UMOD)/Tamm-Horsfall protein, the most abundant human urinary protein, plays a key role in chronic kidney diseases and is a promising therapeutic target for hypertension. Via its bipartite zona pellucida module (ZP-N/ZP-C), UMOD forms extracellular filaments that regulate kidney electrolyte balance and innate immunity, as well as protect against renal stones. Moreover, salt-dependent aggregation of UMOD filaments in the urine generates a soluble molecular net that captures uropathogenic bacteria and facilitates their clearance. Despite the functional importance of its homopolymers, no structural information is available on UMOD and how it self-assembles into filaments. Here, we report the crystal structures of polymerization regions of human UMOD and mouse ZP2, an essential sperm receptor protein that is structurally related to UMOD but forms heteropolymers. The structure of UMOD reveals that an extensive hydrophobic interface mediates ZP-N domain homodimerization. This arrangement is required for filament formation and is directed by an ordered ZP-N/ZP-C linker that is not observed in ZP2 but is conserved in the sequence of deafness/Crohn's disease-associated homopolymeric glycoproteins α-tectorin (TECTA) and glycoprotein 2 (GP2). Our data provide an example of how interdomain linker plasticity can modulate the function of structurally similar multidomain proteins. Moreover, the architecture of UMOD rationalizes numerous pathogenic mutations in both UMOD and TECTA genes. #1: ジャーナル: Proc Soc Exp Biol Med / 年: 1950 タイトル: Characterization and separation of an inhibitor of viral hemagglutination present in urine. #2: ジャーナル: J.Biol.Chem. / 年: 1991 タイトル: Isolation of the cDNA encoding glycoprotein-2 (GP-2), the major zymogen granule membrane protein. Homology to uromodulin/Tamm-Horsfall protein. 著者: Hoops, T.C. / Rindler, M.J. #3: ジャーナル: Nat.Genet. / 年: 1998 タイトル: Mutations in the human alpha-tectorin gene cause autosomal dominant non-syndromic hearing impairment. 著者: Verhoeven, K. / Van Laer, L. / Kirschhofer, K. / Legan, P.K. / Hughes, D.C. / Schatteman, I. / Verstreken, M. / Van Hauwe, P. / Coucke, P. / Chen, A. / Smith, R.J. / Somers, T. / Offeciers, F. ...著者: Verhoeven, K. / Van Laer, L. / Kirschhofer, K. / Legan, P.K. / Hughes, D.C. / Schatteman, I. / Verstreken, M. / Van Hauwe, P. / Coucke, P. / Chen, A. / Smith, R.J. / Somers, T. / Offeciers, F.E. / Van de Heyning, P. / Richardson, G.P. / Wachtler, F. / Kimberling, W.J. / Willems, P.J. / Govaerts, P.J. / Van Camp, G. #4: ジャーナル: Nat Cell Biol / 年: 2002 タイトル: The ZP domain is a conserved module for polymerization of extracellular proteins. 著者: Luca Jovine / Huayu Qi / Zev Williams / Eveline Litscher / Paul M Wassarman /  要旨: Many eukaryotic extracellular proteins share a sequence of unknown function, called the zona pellucida (ZP) domain. Among these proteins are the mammalian sperm receptors ZP2 and ZP3, non-mammalian ...Many eukaryotic extracellular proteins share a sequence of unknown function, called the zona pellucida (ZP) domain. Among these proteins are the mammalian sperm receptors ZP2 and ZP3, non-mammalian egg coat proteins, Tamm-Horsfall protein (THP), glycoprotein-2 (GP-2), alpha- and beta-tectorins, transforming growth factor (TGF)-beta receptor III and endoglin, DMBT-1 (deleted in malignant brain tumour-1), NompA (no-mechanoreceptor-potential-A), Dumpy and cuticlin-1 (refs 1,2). Here, we report that the ZP domain of ZP2, ZP3 and THP is responsible for polymerization of these proteins into filaments of similar supramolecular structure. Most ZP domain proteins are synthesized as precursors with carboxy-terminal transmembrane domains or glycosyl phosphatidylinositol (GPI) anchors. Our results demonstrate that the C-terminal transmembrane domain and short cytoplasmic tail of ZP2 and ZP3 are not required for secretion, but are essential for assembly. Finally, we suggest a molecular basis for dominant human hearing disorders caused by point mutations within the ZP domain of alpha-tectorin. #5: ジャーナル: Am J Kidney Dis / 年: 2003 タイトル: Tamm-Horsfall glycoprotein: biology and clinical relevance. 著者: Franca Serafini-Cessi / Nadia Malagolini / Daniela Cavallone / 要旨: Tamm-Horsfall glycoprotein (THP) is the most abundant urinary protein in mammals. Urinary excretion occurs by proteolytic cleavage of the large ectodomain of the glycosyl phosphatidylinositol- ...Tamm-Horsfall glycoprotein (THP) is the most abundant urinary protein in mammals. Urinary excretion occurs by proteolytic cleavage of the large ectodomain of the glycosyl phosphatidylinositol-anchored counterpart exposed at the luminal cell surface of the thick ascending limb of Henle's loop. We describe the physical-chemical structure of human THP and its biosynthesis and interaction with other proteins and leukocytes. The clinical relevance of THP reported here includes: (1) involvement in the pathogenesis of cast nephropathy, urolithiasis, and tubulointerstitial nephritis; (2) abnormalities in urinary excretion in renal diseases; and (3) the recent finding that familial juvenile hyperuricemic nephropathy and autosomal dominant medullary cystic kidney disease 2 arise from mutations of the THP gene. We critically examine the literature on the physiological role and mechanism(s) that promote urinary excretion of THP. Some lines of research deal with the in vitro immunoregulatory activity of THP, termed uromodulin when isolated from urine of pregnant women. However, an immunoregulatory function in vivo has not yet been established. In the most recent literature, there is renewed interest in the capacity of urinary THP to compete efficiently with urothelial cell receptors, such as uroplakins, in adhering to type 1 fimbriated Escherichia coli. This property supports the notion that abundant THP excretion in urine is promoted in the host by selective pressure to obtain an efficient defense against urinary tract infections caused by uropathogenic bacteria. #6: ジャーナル: Proc Natl Acad Sci U S A / 年: 2004 タイトル: A duplicated motif controls assembly of zona pellucida domain proteins. 著者: Luca Jovine / Huayu Qi / Zev Williams / Eveline S Litscher / Paul M Wassarman / 要旨: Many secreted eukaryotic glycoproteins that play fundamental roles in development, hearing, immunity, and cancer polymerize into filaments and extracellular matrices through zona pellucida (ZP) ...Many secreted eukaryotic glycoproteins that play fundamental roles in development, hearing, immunity, and cancer polymerize into filaments and extracellular matrices through zona pellucida (ZP) domains. ZP domain proteins are synthesized as precursors containing C-terminal propeptides that are cleaved at conserved sites. However, the consequences of this processing and the mechanism by which nascent proteins assemble are unclear. By microinjection of mutated DNA constructs into growing oocytes and mammalian cell transfection, we have identified a conserved duplicated motif [EHP (external hydrophobic patch)/IHP (internal hydrophobic patch)] regulating the assembly of mouse ZP proteins. Whereas the transmembrane domain (TMD) of ZP3 can be functionally replaced by an unrelated TMD, mutations in either EHP or IHP do not hinder secretion of full-length ZP3 but completely abolish its assembly. Because mutants truncated before the TMD are not processed, we conclude that the conserved TMD of mammalian ZP proteins does not engage them in specific interactions but is essential for C-terminal processing. Cleavage of ZP precursors results in loss of the EHP, thereby activating secreted polypeptides to assemble by using the IHP within the ZP domain. Taken together, these findings suggest a general mechanism for assembly of ZP domain proteins. #7: ジャーナル: Annu Rev Biochem / 年: 2005 タイトル: Zona pellucida domain proteins. 著者: Luca Jovine / Costel C Darie / Eveline S Litscher / Paul M Wassarman / 要旨: Many eukaryotic proteins share a sequence designated as the zona pellucida (ZP) domain. This structural element, present in extracellular proteins from a wide variety of organisms, from nematodes to ...Many eukaryotic proteins share a sequence designated as the zona pellucida (ZP) domain. This structural element, present in extracellular proteins from a wide variety of organisms, from nematodes to mammals, consists of approximately 260 amino acids with eight conserved cysteine (Cys) residues and is located close to the C terminus of the polypeptide. ZP domain proteins are often glycosylated, modular structures consisting of multiple types of domains. Predictions can be made about some of the structural features of the ZP domain and ZP domain proteins. The functions of ZP domain proteins vary tremendously, from serving as structural components of egg coats, appendicularian mucous houses, and nematode dauer larvae, to serving as mechanotransducers in flies and receptors in mammals and nonmammals. Generally, ZP domain proteins are present in filaments and/or matrices, which is consistent with the role of the domain in protein polymerization. A general mechanism for assembly of ZP domain proteins has been presented. It is likely that the ZP domain plays a common role despite its presence in proteins of widely diverse functions. #8: ジャーナル: Cell / 年: 2010タイトル: Insights into egg coat assembly and egg-sperm interaction from the X-ray structure of full-length ZP3. 著者: Ling Han / Magnus Monné / Hiroki Okumura / Thomas Schwend / Amy L Cherry / David Flot / Tsukasa Matsuda / Luca Jovine / 要旨: ZP3, a major component of the zona pellucida (ZP) matrix coating mammalian eggs, is essential for fertilization by acting as sperm receptor. By retaining a propeptide that contains a polymerization- ...ZP3, a major component of the zona pellucida (ZP) matrix coating mammalian eggs, is essential for fertilization by acting as sperm receptor. By retaining a propeptide that contains a polymerization-blocking external hydrophobic patch (EHP), we determined the crystal structure of an avian homolog of ZP3 at 2.0 Å resolution. The structure unveils the fold of a complete ZP domain module in a homodimeric arrangement required for secretion and reveals how EHP prevents premature incorporation of ZP3 into the ZP. This suggests mechanisms underlying polymerization and how local structural differences, reflected by alternative disulfide patterns, control the specificity of ZP subunit interaction. Close relative positioning of a conserved O-glycan important for sperm binding and the hypervariable, positively selected C-terminal region of ZP3 suggests a concerted role in the regulation of species-restricted gamete recognition. Alternative conformations of the area around the O-glycan indicate how sperm binding could trigger downstream events via intramolecular signaling. #9: ジャーナル: Kidney Int / 年: 2011 タイトル: The rediscovery of uromodulin (Tamm-Horsfall protein): from tubulointerstitial nephropathy to chronic kidney disease. 著者: Luca Rampoldi / Francesco Scolari / Antonio Amoroso / Gianmarco Ghiggeri / Olivier Devuyst / 要旨: Uromodulin (Tamm-Horsfall protein) is the most abundant protein excreted in the urine under physiological conditions. It is exclusively produced in the kidney and secreted into the urine via ...Uromodulin (Tamm-Horsfall protein) is the most abundant protein excreted in the urine under physiological conditions. It is exclusively produced in the kidney and secreted into the urine via proteolytic cleavage. Its biological function is still not fully understood. Uromodulin has been linked to water/electrolyte balance and to kidney innate immunity. Also, studies in knockout mice demonstrated that it has a protective role against urinary tract infections and renal stone formation. Mutations in the gene encoding uromodulin lead to rare autosomal dominant diseases, collectively referred to as uromodulin-associated kidney diseases. They are characterized by progressive tubulointerstitial damage, impaired urinary concentrating ability, hyperuricemia, renal cysts, and progressive renal failure. Novel in vivo studies point at intracellular accumulation of mutant uromodulin as a key primary event in the disease pathogenesis. Recently, genome-wide association studies identified uromodulin as a risk factor for chronic kidney disease (CKD) and hypertension, and suggested that the level of uromodulin in the urine could represent a useful biomarker for the development of CKD. In this review, we summarize these recent investigations, ranging from invalidation studies in mouse to Mendelian disorders and genome-wide associations, which led to a rediscovery of uromodulin and boosted the scientific and clinical interest for this long discovered molecule. | ||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4wrn.cif.gz | 752.9 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4wrn.ent.gz | 637 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 4wrn.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 4wrn_validation.pdf.gz | 1.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 4wrn_full_validation.pdf.gz | 1.1 MB | 表示 | |
| XML形式データ | 4wrn_validation.xml.gz | 45.7 KB | 表示 | |
| CIF形式データ | 4wrn_validation.cif.gz | 62.3 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/wr/4wrnftp://data.pdbj.org/pub/pdb/validation_reports/wr/4wrn | HTTPS FTP |
-関連構造データ









-リンク
 PDBj
PDBj








- 集合体
集合体
| 登録構造単位 | 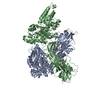
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| 単位格子 |
|
-要素
| #1: タンパク質 | 分子量: 76539.414 Da / 分子数: 2 変異: I28T, D108A, K109A, E198A, N199A, A241H, K245H, K265A, A338V, I343V, E385A, E388A, D389A, R393N,R586A, R588A,I28T, D108A, K109A, E198A, N199A, A241H, K245H, K265A, A338V, I343V, E385A, E388A, ...変異: I28T, D108A, K109A, E198A, N199A, A241H, K245H, K265A, A338V, I343V, E385A, E388A, D389A, R393N,R586A, R588A,I28T, D108A, K109A, E198A, N199A, A241H, K245H, K265A, A338V, I343V, E385A, E388A, D389A, R393N,R586A, R588A 由来タイプ: 組換発現 詳細: THIS PROTEIN IS A CHIMERA. RESIDUES 25-391 ARE FROM E. COLI MALTOSE BINDING PROTEIN (MBP), CORRESPOND TO RESIDUES 27-393 OF SWISS-PROT DATABASE ENTRY P0AEX9 AND CONTAIN MUTATIONS I26T, D106A, ...詳細: THIS PROTEIN IS A CHIMERA. RESIDUES 25-391 ARE FROM E. COLI MALTOSE BINDING PROTEIN (MBP), CORRESPOND TO RESIDUES 27-393 OF SWISS-PROT DATABASE ENTRY P0AEX9 AND CONTAIN MUTATIONS I26T, D106A, K107A, E196A, N197A, A239H, K243H, K263A, A336V, I341V, E383A, E386A, D387A AND R391N (CORRESPONDING TO I28T, D108A, K109A, E198A, N199A, A241H, K245H, K265A, A338V, I343V, E385A, E388A, D389A AND R393N IN P0AEX9). RESIDUES 395-710 ARE FROM HUMAN UROMODULIN PROTEIN AND CORRESPOND TO RESIDUES 295-610 OF SWISS-PROT DATABASE ENTRY P07911 AND CONTAIN MUTATIONS N613Q, R686A AND R688A (CORRESPONDING TO N513Q, R586A AND R588A). 由来: (組換発現) Homo sapiens (ヒト)遺伝子: malE, Z5632, ECs5017, UMOD / プラスミド: pHLSEC / 細胞株 (発現宿主): HEK293S / 発現宿主: Homo sapiens (ヒト)参照: UniProt: P0AEY0, UniProt: P07911, UniProt: P0AEX9*PLUS #2: 多糖 | 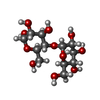  #3: 糖 | 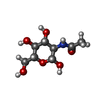  タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C8H15NO6 タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C8H15NO6#4: 化合物 |   分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: ZnHas protein modification | Y | |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 4.91 Å3/Da / 溶媒含有率: 75 % / 解説: Rhombus |
|---|---|
| 結晶化 | 温度: 293 K / 手法: 蒸気拡散法, ハンギングドロップ法 / pH: 8.2 / 詳細: Sodium/potassium tartrate, Tris-HCl / PH範囲: 7.0 - 8.4 |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: ESRF / ビームライン: ID29 / 波長: 0.97625 Å |
| 検出器 | タイプ: DECTRIS PILATUS 6M-F / 検出器: PIXEL / 日付: 2013年1月24日 |
| 放射 | モノクロメーター: Si Crystal / プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.97625 Å / 相対比: 1 |
| 反射 | 解像度: 3.2→35 Å / Num. all: 47837 / Num. obs: 47837 / % possible obs: 99.41 % / 冗長度: 6.3 % / Biso Wilson estimate: 108.3 Å2 / Rmerge(I) obs: 0.08846 / Net I/σ(I): 16.21 |
| 反射 シェル | 解像度: 3.2→3.314 Å / 冗長度: 6.2 % / Rmerge(I) obs: 1.462 / Mean I/σ(I) obs: 1.29 / % possible all: 99.52 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 3SEX 解像度: 3.2→34.976 Å / SU ML: 0.49 / 交差検証法: FREE R-VALUE / σ(F): 1.33 / 位相誤差: 28.21 / 立体化学のターゲット値: ML
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 141.3 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 3.2→34.976 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
