Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4v75: E. coli 70S-fMetVal-tRNAVal-tRNAfMet complex in classic post-tran... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4v75 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | E. coli 70S-fMetVal-tRNAVal-tRNAfMet complex in classic post-translocation state (post1) | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | RIBOSOME / cryo-EM refinement / tRNA / translocation intermediate | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationstringent response / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / misfolded RNA binding / Group I intron splicing / RNA folding / translational termination / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis ...stringent response / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / misfolded RNA binding / Group I intron splicing / RNA folding / translational termination / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis / RNA-binding transcription regulator activity / four-way junction DNA binding / negative regulation of cytoplasmic translation / DnaA-L2 complex / regulation of mRNA stability / translation repressor activity / negative regulation of translational initiation / negative regulation of DNA-templated DNA replication initiation / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing / regulation of DNA-templated transcription elongation / transcription elongation factor complex / response to reactive oxygen species / cytosolic ribosome assembly / ribosome assembly / assembly of large subunit precursor of preribosome / transcription antitermination / DNA endonuclease activity / regulation of cell growth / DNA-templated transcription termination / response to radiation / maintenance of translational fidelity / mRNA 5'-UTR binding / regulation of translation / large ribosomal subunit / transferase activity / ribosomal small subunit assembly / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / 5S rRNA binding / ribosomal large subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / response to antibiotic / negative regulation of DNA-templated transcription / hydrolase activity / mRNA binding / DNA binding / RNA binding / zinc ion binding / membrane / cytoplasm / cytosol Similarity search - Function | ||||||||||||
| Biological species |  | ||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 12 Å | ||||||||||||
 Authors Authors | Blau, C. / Bock, L.V. / Schroder, G.F. / Davydov, I. / Fischer, N. / Stark, H. / Rodnina, M.V. / Vaiana, A.C. / Grubmuller, H. | ||||||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2013 Title: Energy barriers and driving forces in tRNA translocation through the ribosome. Authors: Lars V Bock / Christian Blau / Gunnar F Schröder / Iakov I Davydov / Niels Fischer / Holger Stark / Marina V Rodnina / Andrea C Vaiana / Helmut Grubmüller /  Abstract: During protein synthesis, tRNAs move from the ribosome's aminoacyl to peptidyl to exit sites. Here we investigate conformational motions during spontaneous translocation, using molecular dynamics ...During protein synthesis, tRNAs move from the ribosome's aminoacyl to peptidyl to exit sites. Here we investigate conformational motions during spontaneous translocation, using molecular dynamics simulations of 13 intermediate-translocation-state models obtained by combining Escherichia coli ribosome crystal structures with cryo-EM data. Resolving fast transitions between states, we find that tRNA motions govern the transition rates within the pre- and post-translocation states. Intersubunit rotations and L1-stalk motion exhibit fast intrinsic submicrosecond dynamics. The L1 stalk drives the tRNA from the peptidyl site and links intersubunit rotation to translocation. Displacement of tRNAs is controlled by 'sliding' and 'stepping' mechanisms involving conserved L16, L5 and L1 residues, thus ensuring binding to the ribosome despite large-scale tRNA movement. Our results complement structural data with a time axis, intrinsic transition rates and molecular forces, revealing correlated functional motions inaccessible by other means. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4v75.cif.gz | 3.3 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4v75.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 4v75.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/v7/4v75ftp://data.pdbj.org/pub/pdb/validation_reports/v7/4v75 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  1721M  2472C  2473C  2474C  2475C  4v6yC  4v6zC  4v70C  4v71C  4v72C  4v73C  4v74C  4v76C  4v77C  4v78C  4v79C  4v7aC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |

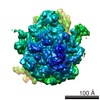
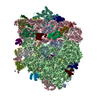

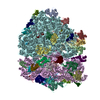

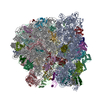



-Links
 PDBj
PDBj






























- Assembly
Assembly
| Deposited unit | 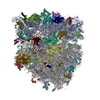
|
|---|---|
| 1 |
|
-Components
-30S ribosomal protein ... , 20 types, 20 molecules ABACADAEAFAGAHAIAJAKALAMANAOAPAQARASATAU
| #1: Protein | Mass: 24277.992 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #2: Protein | Mass: 23207.990 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #3: Protein | Mass: 23514.199 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #4: Protein | Mass: 15828.328 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #5: Protein | Mass: 11667.380 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #6: Protein | Mass: 16788.453 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #7: Protein | Mass: 14146.557 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #8: Protein | Mass: 14580.919 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #9: Protein | Mass: 11221.034 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #10: Protein | Mass: 12513.237 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #11: Protein | Mass: 13768.157 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #12: Protein | Mass: 12657.843 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #13: Protein | Mass: 11606.560 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #14: Protein | Mass: 10290.816 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #15: Protein | Mass: 9063.426 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #16: Protein | Mass: 9287.992 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #17: Protein | Mass: 6490.523 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #18: Protein | Mass: 9081.672 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #19: Protein | Mass: 9532.228 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #20: Protein | Mass: 6091.125 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-RNA chain , 6 types, 6 molecules AAA1A2A3BABB
| #21: RNA chain | Mass: 496892.375 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #22: RNA chain | Mass: 24617.768 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #23: RNA chain | Mass: 4693.746 Da / Num. of mol.: 1 / Source method: obtained synthetically / Details: mRNA |
| #24: RNA chain | Mass: 24848.918 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #54: RNA chain | Mass: 941306.188 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #55: RNA chain | Mass: 38177.762 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
+50S ribosomal protein ... , 30 types, 30 molecules BCBDBEBFBGBHBIBJBKBLBMBNBOBPBQBRBSBTBUBVBWBXBYBZB0B1B2B3B4B5
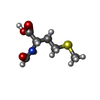
-Non-polymers , 2 types, 2 molecules 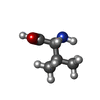
| #57: Chemical | ChemComp-VAL /  Type: L-peptide linking / Mass: 117.146 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C5H11NO2 Type: L-peptide linking / Mass: 117.146 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C5H11NO2 |
|---|---|
| #58: Chemical | ChemComp-FME /  Type: L-peptide linking / Mass: 177.221 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H11NO3S Type: L-peptide linking / Mass: 177.221 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H11NO3S |
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: E. coli 70S-fMetVal-tRNAVal-tRNAfMet complex / Type: RIBOSOME |
|---|---|
| Buffer solution | Name: 50 mM Tris-HCl, 70 mM NH4Cl, 30 mM KCl, 7 mM MgCl2, 0.6 mM spermine, 0.4 mM spermidine pH: 7.5 Details: 50 mM Tris-HCl, 70 mM NH4Cl, 30 mM KCl, 7 mM MgCl2, 0.6 mM spermine, 0.4 mM spermidine |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE / Humidity: 75 % Details: Manual blotting for about 2 seconds prior to plunging into liquid ethane (custom-built CEVS vitrification instrument with dew point temperature adjusted to 18 degrees C) Method: Manual blotting for about 2 seconds Time resolved state: Samples were vitrified at different time points along the reaction coordinate (1, 2, 5 and 20 minutes after addition of deacylated tRNAfMet to 70S-fMetVal-tRNAVal complexes). |
- Electron microscopy imaging
Electron microscopy imaging
| Microscopy | Model: FEI/PHILIPS CM200FEG / Date: May 11, 2008 Details: Objective lens astigmatism was corrected at 200,000 times magnification |
|---|---|
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 160 kV / Illumination mode: SPOT SCAN FIELD EMISSION GUN / Accelerating voltage: 160 kV / Illumination mode: SPOT SCAN |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 161000 X / Calibrated magnification: 162740 X / Nominal defocus max: 2000 nm / Nominal defocus min: 500 nm / Cs: 2 mm |
| Specimen holder | Specimen holder model: GATAN LIQUID NITROGEN / Specimen holder type: Eucentric / Temperature: 77 K |
| Image recording | Electron dose: 20 e/Å2 / Film or detector model: GENERIC TVIPS (4k x 4k) / Details: 4k CCD camera (TVIPS) |
- Processing
Processing
| EM software |
| |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: local | |||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | |||||||||||||||||||||||||||||||||||
| 3D reconstruction | Method: Projection matching / Resolution: 12 Å / Resolution method: FSC 0.5 CUT-OFF / Num. of particles: 13207 / Nominal pixel size: 2.8 Å / Actual pixel size: 2.8 Å Details: Final maps were calculated from 13 datasets acquired at different time points, computationally sorted into distinct substates. Symmetry type: POINT | |||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: Cross-correlation coefficient Details: METHOD--Cross-correlation gradient based REFINEMENT PROTOCOL--flexible fitting, dynamic elastic network | |||||||||||||||||||||||||||||||||||
| Atomic model building |
|