 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 2c1t | ||||||
|---|---|---|---|---|---|---|---|
| Title | Structure of the Kap60p:Nup2 complex | ||||||
 Components Components |
| ||||||
 Keywords Keywords | PROTEIN TRANSPORT/MEMBRANE PROTEIN / ARMADILLO REPEAT / KARYOPHERIN RECYCLING / NLS RELEASE / NUCEAR IMPORT / NUCLEAR PROTEIN / NUCLEOPORIN / PROTEIN TRANSPORT / NUCLEAR TRANSPORT-COMPLEX / NUCLEAR PORE COMPLEX / PHOSPHORYLATION / TRANSLOCATION / PROTEIN TRANSPORT-MEMBRANE PROTEIN complex | ||||||
| Function / homology |  Function and homology information Function and homology informationproteasome localization / mRNA export from nucleus in response to heat stress / transcription-dependent tethering of RNA polymerase II gene DNA at nuclear periphery / protein localization to nuclear inner membrane / Regulation of Glucokinase by Glucokinase Regulatory Protein / : / nuclear pore cytoplasmic filaments / import into nucleus / Regulation of HSF1-mediated heat shock response / post-transcriptional tethering of RNA polymerase II gene DNA at nuclear periphery ...proteasome localization / mRNA export from nucleus in response to heat stress / transcription-dependent tethering of RNA polymerase II gene DNA at nuclear periphery / protein localization to nuclear inner membrane / Regulation of Glucokinase by Glucokinase Regulatory Protein / : / nuclear pore cytoplasmic filaments / import into nucleus / Regulation of HSF1-mediated heat shock response / post-transcriptional tethering of RNA polymerase II gene DNA at nuclear periphery / nuclear pore nuclear basket / importin-alpha family protein binding / NLS-dependent protein nuclear import complex / SUMOylation of SUMOylation proteins / structural constituent of nuclear pore / SUMOylation of RNA binding proteins / silent mating-type cassette heterochromatin formation / SUMOylation of chromatin organization proteins / nuclear import signal receptor activity / protein targeting to membrane / nuclear localization sequence binding / nucleocytoplasmic transport / NLS-bearing protein import into nucleus / poly(A)+ mRNA export from nucleus / nuclear pore / subtelomeric heterochromatin formation / protein export from nucleus / small GTPase binding / disordered domain specific binding / protein import into nucleus / nuclear envelope / nuclear membrane / chromosome, telomeric region / protein-containing complex binding / perinuclear region of cytoplasm / protein-containing complex / mitochondrion / nucleoplasm / nucleus / cytoplasm / cytosol Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.6 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.6 Å | ||||||
 Authors Authors | Matsuura, Y. / Stewart, M. | ||||||
 Citation Citation | Journal: Embo J. / Year: 2005 Title: Nup50/Npap60 Function in Nuclear Import Complex Disassembly and Importin Recycling Authors: Matsuura, Y. / Stewart, M. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 2c1t.cif.gz | 190.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb2c1t.ent.gz | 151.9 KB | Display | PDB format |
| PDBx/mmJSON format | 2c1t.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/c1/2c1tftp://data.pdbj.org/pub/pdb/validation_reports/c1/2c1t | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  2c1mC  1ee4S S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |













-Links
 PDBj
PDBj




- Assembly
Assembly
| Deposited unit | 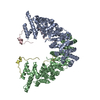
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Unit cell |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS domain:
NCS domain segments:
NCS ensembles :
|
-Components
| #1: Protein | Mass: 50189.902 Da / Num. of mol.: 2 / Fragment: ARM DOMAIN, RESIDUES 88-541 / Mutation: YES Source method: isolated from a genetically manipulated source Source: (gene. exp.) Production host:  #2: Protein | Mass: 5813.649 Da / Num. of mol.: 2 / Fragment: RESIDUES 1-51 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Production host: #3: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 151 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 151 / Source method: isolated from a natural source / Formula: H2OCompound details | IMPORTIN ALPHA SUBUNIT: SPECIFICALLY BINDS TO SUBSTRATES CONTAINING NLS MOTIF AND PROMOTES DOCKING ...IMPORTIN ALPHA SUBUNIT: SPECIFICAL | Sequence details | RESIDUES 1 TO 87 REMOVED. Y397D INTRODUCED | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.4 Å3/Da / Density % sol: 49 % |
|---|---|
| Crystal grow | pH: 6.8 Details: EXPERIMENTAL DETALS GIVEN IN ENTRY 1UN0 AND IN MATSUURA ET AL, EMBO J, 22, 5358, 2003., pH 6.80 |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SRS  / Beamline: PX14.2 / Wavelength: 0.978 / Beamline: PX14.2 / Wavelength: 0.978 |
| Detector | Type: ADSC CCD / Detector: CCD / Date: Apr 21, 2003 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.978 Å / Relative weight: 1 |
| Reflection | Resolution: 2.6→20 Å / Num. obs: 29670 / % possible obs: 97.5 % / Redundancy: 2.3 % / Rmerge(I) obs: 0.09 / Net I/σ(I): 10 |
| Reflection shell | Resolution: 2.6→2.74 Å / Redundancy: 2.3 % / Rmerge(I) obs: 0.53 / Mean I/σ(I) obs: 1.8 / % possible all: 98.3 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRY 1EE4 Resolution: 2.6→20 Å / Cor.coef. Fo:Fc: 0.943 / Cor.coef. Fo:Fc free: 0.914 / SU B: 10.848 / SU ML: 0.229 / Cross valid method: THROUGHOUT / ESU R: 0.673 / ESU R Free: 0.31 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.4 Å / Solvent model: BABINET MODEL WITH MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 49.39 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.6→20 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|