 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-4937 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Cryo-EM 3D map of normal Huntingtin | |||||||||
 マップデータ マップデータ | ||||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | multivalent scaffold platform / PROTEIN BINDING | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報positive regulation of inositol 1,4,5-trisphosphate-sensitive calcium-release channel activity / positive regulation of CAMKK-AMPK signaling cascade / microtubule-based transport / vocal learning / positive regulation of mitophagy / regulation of CAMKK-AMPK signaling cascade / profilin binding / positive regulation of cilium assembly / retrograde vesicle-mediated transport, Golgi to endoplasmic reticulum / vesicle transport along microtubule ...positive regulation of inositol 1,4,5-trisphosphate-sensitive calcium-release channel activity / positive regulation of CAMKK-AMPK signaling cascade / microtubule-based transport / vocal learning / positive regulation of mitophagy / regulation of CAMKK-AMPK signaling cascade / profilin binding / positive regulation of cilium assembly / retrograde vesicle-mediated transport, Golgi to endoplasmic reticulum / vesicle transport along microtubule / positive regulation of aggrephagy / positive regulation of lipophagy / Golgi organization / dynein intermediate chain binding / establishment of mitotic spindle orientation / dynactin binding / phosphoprotein phosphatase activity / Regulation of MECP2 expression and activity / postsynaptic cytosol / beta-tubulin binding / presynaptic cytosol / inclusion body / heat shock protein binding / centriole / autophagosome / cytoplasmic vesicle membrane / negative regulation of extrinsic apoptotic signaling pathway / protein destabilization / kinase binding / p53 binding / late endosome / transmembrane transporter binding / early endosome / positive regulation of apoptotic process / axon / apoptotic process / dendrite / perinuclear region of cytoplasm / endoplasmic reticulum / Golgi apparatus / protein-containing complex / nucleoplasm / identical protein binding / nucleus / cytosol / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 9.6 Å | |||||||||
 データ登録者 データ登録者 | Jung T / Tamo G | |||||||||
| 資金援助 |  韓国, 2件 韓国, 2件
| |||||||||
 引用 引用 | ジャーナル: Structure / 年: 2020 タイトル: The Polyglutamine Expansion at the N-Terminal of Huntingtin Protein Modulates the Dynamic Configuration and Phosphorylation of the C-Terminal HEAT Domain. 著者: Taeyang Jung / Baehyun Shin / Giorgio Tamo / Hyeongju Kim / Ravi Vijayvargia / Alexander Leitner / Maria J Marcaida / Juan Astorga-Wells / Roy Jung / Ruedi Aebersold / Matteo Dal Peraro / ...著者: Taeyang Jung / Baehyun Shin / Giorgio Tamo / Hyeongju Kim / Ravi Vijayvargia / Alexander Leitner / Maria J Marcaida / Juan Astorga-Wells / Roy Jung / Ruedi Aebersold / Matteo Dal Peraro / Hans Hebert / Ihn Sik Seong / Ji-Joon Song /    要旨: The polyQ expansion in huntingtin protein (HTT) is the prime cause of Huntington's disease (HD). The recent cryoelectron microscopy (cryo-EM) structure of HTT-HAP40 complex provided the structural ...The polyQ expansion in huntingtin protein (HTT) is the prime cause of Huntington's disease (HD). The recent cryoelectron microscopy (cryo-EM) structure of HTT-HAP40 complex provided the structural information on its HEAT-repeat domains. Here, we present analyses of the impact of polyQ length on the structure and function of HTT via an integrative structural and biochemical approach. The cryo-EM analysis of normal (Q23) and disease (Q78) type HTTs shows that the structures of apo HTTs significantly differ from the structure of HTT in a HAP40 complex and that the polyQ expansion induces global structural changes in the relative movements among the HTT domains. In addition, we show that the polyQ expansion alters the phosphorylation pattern across HTT and that Ser2116 phosphorylation in turn affects the global structure and function of HTT. These results provide a molecular basis for the effect of the polyQ segment on HTT structure and activity, which may be important for HTT pathology. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_4937.map.gz | 55.2 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-4937-v30.xmlemd-4937.xml | 17.1 KB 17.1 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_4937_fsc.xml | 10.5 KB | 表示 | FSCデータファイル |
| 画像 |  emd_4937.png emd_4937.png | 43.4 KB | ||
| Filedesc metadata | emd-4937.cif.gz | 8 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-4937ftp://ftp.pdbj.org/pub/emdb/structures/EMD-4937 http://ftp.pdbj.org/pub/emdb/structures/EMD-4937ftp://ftp.pdbj.org/pub/emdb/structures/EMD-4937 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_4937_validation.pdf.gz | 413.2 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_4937_full_validation.pdf.gz | 412.8 KB | 表示 | |
| XML形式データ | emd_4937_validation.xml.gz | 10.7 KB | 表示 | |
| CIF形式データ | emd_4937_validation.cif.gz | 14 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-4937ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-4937 | HTTPS FTP |
-関連構造データ









-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |




-マップ
| ファイル | ダウンロード / ファイル: emd_4937.map.gz / 形式: CCP4 / 大きさ: 59.6 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.06 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
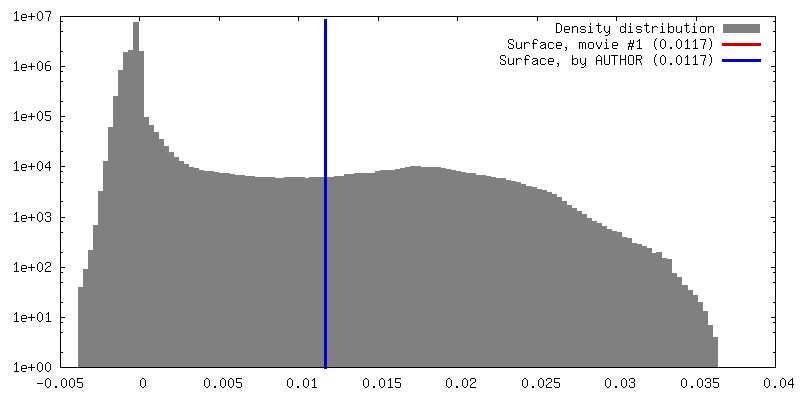
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)




















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : normal-type human huntingtin
| 全体 | 名称: normal-type human huntingtin |
|---|---|
| 要素 |
|
-超分子 #1: normal-type human huntingtin
| 超分子 | 名称: normal-type human huntingtin / タイプ: organelle_or_cellular_component / ID: 1 / 親要素: 0 / 含まれる分子: all |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 350 KDa |
-分子 #1: Huntingtin
| 分子 | 名称: Huntingtin / タイプ: protein_or_peptide / ID: 1 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 347.974906 KDa |
| 組換発現 | 生物種:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ) |
| 配列 | 文字列: MATLEKLMKA FESLKSFQQQ QQQQQQQQQQ QQQQQQQQPP PPPPPPPPPQ LPQPPPQAQP LLPQPQPPPP PPPPPPGPAV AEEPLHRPK KELSATKKDR VNHCLTICEN IVAQSVRNSP EFQKLLGIAM ELFLLCSDDA ESDVRMVADE CLNKVIKALM D SNLPRLQL ...文字列: MATLEKLMKA FESLKSFQQQ QQQQQQQQQQ QQQQQQQQPP PPPPPPPPPQ LPQPPPQAQP LLPQPQPPPP PPPPPPGPAV AEEPLHRPK KELSATKKDR VNHCLTICEN IVAQSVRNSP EFQKLLGIAM ELFLLCSDDA ESDVRMVADE CLNKVIKALM D SNLPRLQL ELYKEIKKNG APRSLRAALW RFAELAHLVR PQKCRPYLVN LLPCLTRTSK RPEESVQETL AAAVPKIMAS FG NFANDNE IKVLLKAFIA NLKSSSPTIR RTAAGSAVSI CQHSRRTQYF YSWLLNVLLG LLVPVEDEHS TLLILGVLLT LRY LVPLLQ QQVKDTSLKG SFGVTRKEME VSPSAEQLVQ VYELTLHHTQ HQDHNVVTGA LELLQQLFRT PPPELLQTLT AVGG IGQLT AAKEESGGRS RSGSIVELIA GGGSSCSPVL SRKQKGKVLL GEEEALEDDS ESRSDVSSSA LTASVKDEIS GELAA SSGV STPGSAGHDI ITEQPRSQHT LQADSVDLAS CDLTSSATDG DEEDILSHSS SQVSAVPSDP AMDLNDGTQA SSPISD SSQ TTTEGPDSAV TPSDSSEIVL DGTDNQYLGL QIGQPQDEDE EATGILPDEA SEAFRNSSMA LQQAHLLKNM SHCRQPS DS SVDKFVLRDE ATEPGDQENK PCRIKGDIGQ STDDDSAPLV HCVRLLSASF LLTGGKNVLV PDRDVRVSVK ALALSCVG A AVALHPESFF SKLYKVPLDT TEYPEEQYVS DILNYIDHGD PQVRGATAIL CGTLICSILS RSRFHVGDWM GTIRTLTGN TFSLADCIPL LRKTLKDESS VTCKLACTAV RNCVMSLCSS SYSELGLQLI IDVLTLRNSS YWLVRTELLE TLAEIDFRLV SFLEAKAEN LHRGAHHYTG LLKLQERVLN NVVIHLLGDE DPRVRHVAAA SLIRLVPKLF YKCDQGQADP VVAVARDQSS V YLKLLMHE TQPPSHFSVS TITRIYRGYN LLPSITDVTM ENNLSRVIAA VSHELITSTT RALTFGCCEA LCLLSTAFPV CI WSLGWHC GVPPLSASDE SRKSCTVGMA TMILTLLSSA WFPLDLSAHQ DALILAGNLL AASAPKSLRS SWASEEEANP AAT KQEEVW PALGDRALVP MVEQLFSHLL KVINICAHVL DDVAPGPAIK AALPSLTNPP SLSPIRRKGK EKEPGEQASV PLSP KKGSE ASAASRQSDT SGPVTTSKSS SLGSFYHLPS YLRLHDVLKA THANYKVTLD LQNSTEKFGG FLRSALDVLS QILEL ATLQ DIGKCVEEIL GYLKSCFSRE PMMATVCVQQ LLKTLFGTNL ASQFDGLSSN PSKSQGRAQR LGSSSVRPGL YHYCFM APY THFTQALADA SLRNMVQAEQ ENDTSGWFDV LQKVSTQLKT NLTSVTKNRA DKNAIHNHIR LFEPLVIKAL KQYTTTT CV QLQKQVLDLL AQLVQLRVNY CLLDSDQVFI GFVLKQFEYI EVGQFRESEA IIPNIFFFLV LLSYERYHSK QIIGIPKI I QLCDGIMASG RKAVTHAIPA LQPIVHDLFV LRGTNKADAG KELETQKEVV VSMLLRLIQY HQVLEMFILV LQQCHKENE DKWKRLSRQI ADIILPMLAK QQMHIDSHEA LGVLNTLFEI LAPSSLRPVD MLLRSMFVTP NTMASVSTVQ LWISGILAIL RVLISQSTE DIVLSRIQEL SFSPYLISCT VINRLRDGDS TSTLEEHSEG KQIKNLPEET FSRFLLQLVG ILLEDIVTKQ L KVEMSEQQ HTFYCQELGT LLMCLIHIFK SGMFRRITAA ATRLFRSDGC GGSFYTLDSL NLRARSMITT HPALVLLWCQ IL LLVNHTD YRWWAEVQQT PKRHSLSSTK LLSPQMSGEE EDSDLAAKLG MCNREIVRRG ALILFCDYVC QNLHDSEHLT WLI VNHIQD LISLSHEPPV QDFISAVHRN SAASGLFIQA IQSRCENLST PTMLKKTLQC LEGIHLSQSG AVLTLYVDRL LCTP FRVLA RMVDILACRR VEMLLAANLQ SSMAQLPMEE LNRIQEYLQS SGLAQRHQRL YSLLDRFRLS TMQDSLSPSP PVSSH PLDG DGHVSLETVS PDKDWYVHLV KSQCWTRSDS ALLEGAELVN RIPAEDMNAF MMNSEFNLSL LAPCLSLGMS EISGGQ KSA LFEAAREVTL ARVSGTVQQL PAVHHVFQPE LPAEPAAYWS KLNDLFGDAA LYQSLPTLAR ALAQYLVVVS KLPSHLH LP PEKEKDIVKF VVATLEALSW HLIHEQIPLS LDLQAGLDCC CLALQLPGLW SVVSSTEFVT HACSLIHCVH FILEAVAV Q PGEQLLSPER RTNTPKAISE EEEEVDPNTQ NPKYITAACE MVAEMVESLQ SVLALGHKRN SGVPAFLTPL LRNIIISLA RLPLVNSYTR VPPLVWKLGW SPKPGGDFGT AFPEIPVEFL QEKEVFKEFI YRINTLGWTS RTQFEETWAT LLGVLVTQPL VMEQEESPP EEDTERTQIN VLAVQAITSL VLSAMTVPVA GNPAVSCLEQ QPRNKPLKAL DTRFGRKLSI IRGIVEQEIQ A MVSKRENI ATHHLYQAWD PVPSLSPATT GALISHEKLL LQINPERELG SMSYKLGQVS IHSVWLGNSI TPLREEEWDE EE EEEADAP APSSPPTSPV NSRKHRAGVD IHSCSQFLLE LYSRWILPSS SARRTPAILI SEVVRSLLVV SDLFTERNQF ELM YVTLTE LRRVHPSEDE ILAQYLVPAT CKAAAVLGMD KAVAEPVSRL LESTLRSSHL PSRVGALHGV LYVLECDLLD DTAK QLIPV ISDYLLSNLK GIAHCVNIHS QQHVLVMCAT AFYLIENYPL DVGPEFSASI IQMCGVMLSG SEESTPSIIY HCALR GLER LLLSEQLSRL DAESLVKLSV DRVNVHSPHR AMAALGLMLT CMYTGKEKVS PGRTSDPNPA APDSESVIVA MERVSV LFD RIRKGFPCEA RVVARILPQF LDDFFPPQDI MNKVIGEFLS NQQPYPQFMA TVVYKVFQTL HSTGQSSMVR DWVMLSL SN FTQRAPVAMA TWSLSCFFVS ASTSPWVAAI LPHVISRMGK LEQVDVNLFC LVATDFYRHQ IEEELDRRAF QSVLEVVA A PGSPYHRLLT CLRNVHKVTT C UniProtKB: Huntingtin |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 0.06 mg/mL | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | pH: 7.5 構成要素:
| |||||||||
| グリッド | モデル: Quantifoil R2/1 / 材質: COPPER / メッシュ: 300 / 支持フィルム - 材質: GRAPHENE OXIDE / 支持フィルム - トポロジー: CONTINUOUS | |||||||||
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 288 K / 装置: FEI VITROBOT MARK I / 詳細: blot 9 seconds incubate 30 seconds. | |||||||||
| 詳細 | HTT was mixed with final 0.05% of Octyl glucoside. |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 特殊光学系 | 位相板: VOLTA PHASE PLATE |
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: COUNTING / デジタル化 - 画像ごとのフレーム数: 2-40 / 実像数: 2331 / 平均露光時間: 8.0 sec. / 平均電子線量: 40.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 最大 デフォーカス(補正後): 1.0 µm / 倍率(補正後): 47170 / 照射モード: SPOT SCAN / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 0.8 µm / 最小 デフォーカス(公称値): 0.4 µm |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |


