 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- EMDB-47721: Human proteasome in resting state conformation bound to TXNL1 in ... -
+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Human proteasome in resting state conformation bound to TXNL1 in Forward conformation | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | 26S Proteasome / Nub1 / Fat10 / MOTOR PROTEIN / HYDROLASE-PROTEIN BINDING complex | |||||||||
| Function / homology |  Function and homology information Function and homology informationregulation of ubiquitin-dependent protein catabolic process / disulfide oxidoreductase activity / thyrotropin-releasing hormone receptor binding / nuclear proteasome complex / host-mediated perturbation of viral transcription / positive regulation of inclusion body assembly / Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / proteasome accessory complex ...regulation of ubiquitin-dependent protein catabolic process / disulfide oxidoreductase activity / thyrotropin-releasing hormone receptor binding / nuclear proteasome complex / host-mediated perturbation of viral transcription / positive regulation of inclusion body assembly / Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / proteasome accessory complex / purine ribonucleoside triphosphate binding / integrator complex / proteasome regulatory particle / CD8-positive, alpha-beta T cell differentiation / thymic T cell selection / CD8-positive, alpha-beta T cell homeostasis / cytosolic proteasome complex / positive regulation of proteasomal protein catabolic process / proteasome-activating activity / Antigen processing: Ub, ATP-independent proteasomal degradation / RND1 GTPase cycle / RND2 GTPase cycle / RND3 GTPase cycle / proteasome regulatory particle, lid subcomplex / proteasome regulatory particle, base subcomplex / RHOBTB1 GTPase cycle / T-helper 1 cell differentiation / negative regulation of regulatory T cell differentiation / cellular response to type I interferon / protein K63-linked deubiquitination / RHOV GTPase cycle / metal-dependent deubiquitinase activity / negative regulation of programmed cell death / Regulation of ornithine decarboxylase (ODC) / proteasome core complex / Proteasome assembly / T-helper 17 cell differentiation / Cross-presentation of soluble exogenous antigens (endosomes) / transcription factor binding / K63-linked deubiquitinase activity / Somitogenesis / Homologous DNA Pairing and Strand Exchange / Defective homologous recombination repair (HRR) due to BRCA1 loss of function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA1 binding function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA2/RAD51/RAD51C binding function / Lewy body / Resolution of D-loop Structures through Synthesis-Dependent Strand Annealing (SDSA) / flagellated sperm motility / Resolution of D-loop Structures through Holliday Junction Intermediates / proteasome binding / RHOU GTPase cycle / Impaired BRCA2 binding to RAD51 / regulation of protein catabolic process / myofibril / protein-disulfide reductase activity / response to tumor necrosis factor / AMPK-induced ERAD and lysosome mediated degradation of PD-L1(CD274) / GSK3B-mediated proteasomal degradation of PD-L1(CD274) / SPOP-mediated proteasomal degradation of PD-L1(CD274) / proteasomal ubiquitin-independent protein catabolic process / positive regulation of RNA polymerase II transcription preinitiation complex assembly / Ribosome Quality Control (RQC) complex extracts and degrades nascent peptide / general transcription initiation factor binding / proteasome storage granule / Presynaptic phase of homologous DNA pairing and strand exchange / protein deubiquitination / polyubiquitin modification-dependent protein binding / NF-kappaB binding / RHOBTB2 GTPase cycle / endopeptidase activator activity / mRNA export from nucleus / proteasome core complex, alpha-subunit complex / proteasome assembly / SARS-CoV-1 targets host intracellular signalling and regulatory pathways / immune system process / regulation of G1/S transition of mitotic cell cycle / enzyme regulator activity / ERAD pathway / ciliary tip / response to type II interferon / positive regulation of interleukin-2 production / inclusion body / regulation of proteasomal protein catabolic process / TBP-class protein binding / : / proteasome complex / stem cell differentiation / sarcomere / proteasomal protein catabolic process / Regulation of activated PAK-2p34 by proteasome mediated degradation / sperm end piece / ubiquitin binding / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / negative regulation of inflammatory response to antigenic stimulus / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Asymmetric localization of PCP proteins / Ubiquitin-dependent degradation of Cyclin D / lipopolysaccharide binding / SCF-beta-TrCP mediated degradation of Emi1 Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 2.87 Å | |||||||||
 Authors Authors | Arkinson C / Gee CL / Martin A | |||||||||
| Funding support |  United States, 1 items United States, 1 items
| |||||||||
 Citation Citation | Journal: bioRxiv / Year: 2024 Title: Structural landscape of AAA+ ATPase motor states in the substrate-degrading human 26S proteasome reveals conformation-specific binding of TXNL1. Authors: Connor Arkinson / Christine L Gee / Zeyuan Zhang / Ken C Dong / Andreas Martin / Abstract: The 26S proteasome targets many cellular proteins for degradation during general homeostasis, protein quality control, and the regulation of vital processes. A broad range of proteasome-interacting ...The 26S proteasome targets many cellular proteins for degradation during general homeostasis, protein quality control, and the regulation of vital processes. A broad range of proteasome-interacting cofactors thereby modulates these functions and aids in substrate degradation. Here, we solved several high-resolution structures of the redox active cofactor TXNL1 bound to the human 26S proteasome at saturating and sub-stoichiometric concentrations by time resolved cryo-EM. We identified distinct binding modes of TXNL1 that depend on the proteasome conformational and ATPase motor states. Together with biophysical and biochemical experiments, our structural studies reveal that the resting-state proteasome prior to substrate engagement with the ATPase motor binds TXNL1 with low affinity and in variable positions on top of the Rpn11 deubiquitinase. In contrast, the actively degrading proteasome shows additional interactions leading to high-affinity TXNL1 binding, whereby TXNL1's C-terminal tail covers the catalytic groove of the Rpn11 deubiquitinase and coordinates the active-site Zn. Furthermore, these cryo-EM structures of the degrading proteasome capture the ATPase hexamer in all registers of spiral-staircase arrangements and thus visualize the complete ATP-hydrolysis cycle of the AAA+ motor, indicating temporally asymmetric hydrolysis and conformational changes in bursts during mechanical substrate unfolding and translocation. Remarkably, we catch the proteasome in the act of unfolding the beta-barrel mEos3.2 substrate while the ATPase hexamer is in a particular spiral staircase register. Our findings challenge current models for protein translocation through hexameric AAA+ motors and reveal how the proteasome uses its distinct but broad range of conformational states to coordinate cofactor binding and substrate processing. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_47721.map.gz | 75.8 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-47721-v30.xmlemd-47721.xml | 52.5 KB 52.5 KB | Display Display | EMDB header |
| Images |  emd_47721.png emd_47721.png | 121 KB | ||
| Filedesc metadata | emd-47721.cif.gz | 14.2 KB | ||
| Others | emd_47721_half_map_1.map.gzemd_47721_half_map_2.map.gz | 139.2 MB 139.2 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-47721ftp://ftp.pdbj.org/pub/emdb/structures/EMD-47721 http://ftp.pdbj.org/pub/emdb/structures/EMD-47721ftp://ftp.pdbj.org/pub/emdb/structures/EMD-47721 | HTTPS FTP |
-Related structure data
| Related structure data |  9e8iMC  9e8gC  9e8hC  9e8jC  9e8kC  9e8lC  9e8nC  9e8oC 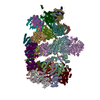 9e8qC  9pdiC  9pdlC  9pdnC  9pf1C M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |














-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |

















-Map
| File | Download / File: emd_47721.map.gz / Format: CCP4 / Size: 149.9 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.048 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
-Half map: #1
| File | emd_47721_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








-Half map: #2
| File | emd_47721_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








- Sample components
Sample components
+Entire : Human 26S proteasome complexed with Nub1, Fat10 and TXNL1
+Supramolecule #1: Human 26S proteasome complexed with Nub1, Fat10 and TXNL1
+Macromolecule #1: 26S proteasome regulatory subunit 7
+Macromolecule #2: 26S proteasome regulatory subunit 4
+Macromolecule #3: 26S protease regulatory subunit 8
+Macromolecule #4: 26S proteasome regulatory subunit 6B
+Macromolecule #5: 26S protease regulatory subunit 10B
+Macromolecule #6: 26S proteasome regulatory subunit 6A
+Macromolecule #7: Proteasome subunit alpha type-6
+Macromolecule #8: Proteasome subunit alpha type-2
+Macromolecule #9: Proteasome subunit alpha type-4
+Macromolecule #10: Proteasome subunit alpha type-7
+Macromolecule #11: Proteasome subunit alpha type-5
+Macromolecule #12: Proteasome subunit alpha type-1
+Macromolecule #13: Proteasome subunit alpha type-3
+Macromolecule #14: 26S proteasome non-ATPase regulatory subunit 1
+Macromolecule #15: 26S proteasome non-ATPase regulatory subunit 3
+Macromolecule #16: 26S proteasome non-ATPase regulatory subunit 12
+Macromolecule #17: 26S proteasome non-ATPase regulatory subunit 11
+Macromolecule #18: 26S proteasome non-ATPase regulatory subunit 6
+Macromolecule #19: 26S proteasome non-ATPase regulatory subunit 7
+Macromolecule #20: 26S proteasome non-ATPase regulatory subunit 13
+Macromolecule #21: 26S proteasome non-ATPase regulatory subunit 4
+Macromolecule #22: 26S proteasome non-ATPase regulatory subunit 14
+Macromolecule #23: 26S proteasome non-ATPase regulatory subunit 8
+Macromolecule #24: 26S proteasome complex subunit SEM1
+Macromolecule #25: 26S proteasome non-ATPase regulatory subunit 2
+Macromolecule #26: Isoform 2 of NEDD8 ultimate buster 1

+Macromolecule #27: Thioredoxin-like protein 1
+Macromolecule #28: ADENOSINE-5'-TRIPHOSPHATE

+Macromolecule #29: MAGNESIUM ION
+Macromolecule #30: ADENOSINE-5'-DIPHOSPHATE

+Macromolecule #31: ZINC ION
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.4 Details: 30 mM HEPES pH7.4, 25 mM NaCl, 25 mM KCl, 3% (v/v) glycerol, 5 mM MgCl2 2 mM ATP and 0.5 mM TCEP. |
|---|---|
| Vitrification | Cryogen name: ETHANE / Chamber humidity: 100 % / Chamber temperature: 285 K / Instrument: FEI VITROBOT MARK IV |
- Electron microscopy
Electron microscopy
| Microscope | TFS KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K3 (6k x 4k) / Average electron dose: 50.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Nominal defocus max: 1.7 µm / Nominal defocus min: 0.5 µm |
| Sample stage | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Cooling holder cryogen: NITROGEN |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
