 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-3339 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
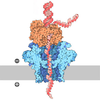
| タイトル | Atomic cryoEM structure of Hsp90/Cdc37/Cdk4 complex | |||||||||
 マップデータ マップデータ | Reconstruction of Hsp90:Cdc37:Cdk4 complex. Part of series of maps, the highest resolution map being EMD-3337. This one is same as EMD-3337, but un-sharpened and filtered to 5.5A. This map has more coherent density for Cdk4 C-lobe. | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | Hsp90 / Cdc37 / Cdk4 / chaperone / kinase / unfolding | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報cyclin D3-CDK4 complex / cyclin D1-CDK4 complex / cyclin D2-CDK4 complex / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 / citrulline metabolic process / regulation of transcription initiation by RNA polymerase II / regulation of type II interferon-mediated signaling pathway / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 ...cyclin D3-CDK4 complex / cyclin D1-CDK4 complex / cyclin D2-CDK4 complex / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 / citrulline metabolic process / regulation of transcription initiation by RNA polymerase II / regulation of type II interferon-mediated signaling pathway / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Drug-mediated inhibition of CDK4/CDK6 activity / regulation of type B pancreatic cell proliferation / HSP90-CDC37 chaperone complex / very long-chain fatty acid metabolic process / negative regulation of proteasomal protein catabolic process / Aryl hydrocarbon receptor signalling / aryl hydrocarbon receptor complex / cellular response to ionomycin / histone methyltransferase binding / Transcriptional regulation by RUNX2 / dynein axonemal particle / cellular response to phorbol 13-acetate 12-myristate / receptor ligand inhibitor activity / : / positive regulation of type 2 mitophagy / positive regulation of protein localization to cell surface / regulation of cyclin-dependent protein serine/threonine kinase activity / ATP-dependent protein binding / protein kinase regulator activity / protein folding chaperone complex / cyclin-dependent protein serine/threonine kinase regulator activity / post-transcriptional regulation of gene expression / Respiratory syncytial virus genome replication / telomerase holoenzyme complex assembly / Uptake and function of diphtheria toxin / positive regulation of transforming growth factor beta receptor signaling pathway / Drug-mediated inhibition of ERBB2 signaling / Resistance of ERBB2 KD mutants to trastuzumab / Resistance of ERBB2 KD mutants to sapitinib / Resistance of ERBB2 KD mutants to tesevatinib / Resistance of ERBB2 KD mutants to neratinib / Resistance of ERBB2 KD mutants to osimertinib / Resistance of ERBB2 KD mutants to afatinib / Resistance of ERBB2 KD mutants to AEE788 / Resistance of ERBB2 KD mutants to lapatinib / Drug resistance in ERBB2 TMD/JMD mutants / PTK6 Regulates Cell Cycle / regulation of type I interferon-mediated signaling pathway / dendritic growth cone / TPR domain binding / Defective binding of RB1 mutants to E2F1,(E2F2, E2F3) / Assembly and release of respiratory syncytial virus (RSV) virions / The NLRP3 inflammasome / protein phosphatase activator activity / Sema3A PAK dependent Axon repulsion / regulation of protein ubiquitination / HSF1-dependent transactivation / bicellular tight junction / positive regulation of G2/M transition of mitotic cell cycle / response to unfolded protein / HSF1 activation / cyclin-dependent kinase / cyclin-dependent protein serine/threonine kinase activity / telomere maintenance via telomerase / Attenuation phase / chaperone-mediated protein complex assembly / protein targeting / axonal growth cone / RHOBTB2 GTPase cycle / Purinergic signaling in leishmaniasis infection / cyclin-dependent protein kinase holoenzyme complex / supramolecular fiber organization / : / regulation of G2/M transition of mitotic cell cycle / DNA polymerase binding / heat shock protein binding / Signaling by ERBB2 / negative regulation of proteasomal ubiquitin-dependent protein catabolic process / protein folding chaperone / peptide binding / cellular response to interleukin-4 / cyclin binding / ESR-mediated signaling / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / Constitutive Signaling by Overexpressed ERBB2 / placenta development / nitric-oxide synthase regulator activity / Ubiquitin-dependent degradation of Cyclin D / positive regulation of cell differentiation / Hsp90 protein binding / ATP-dependent protein folding chaperone / Signaling by ERBB2 TMD/JMD mutants / G1/S transition of mitotic cell cycle / Constitutive Signaling by EGFRvIII / Signaling by ERBB2 ECD mutants / DDX58/IFIH1-mediated induction of interferon-alpha/beta / Oncogene Induced Senescence / Signaling by ERBB2 KD Mutants / Regulation of actin dynamics for phagocytic cup formation / Transcriptional regulation of white adipocyte differentiation 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 5.6 Å | |||||||||
 データ登録者 データ登録者 | Verba KA / Wang RYR / Arakawa A / Liu Y / Shirouzu M / Yokoyama S / Agard DA | |||||||||
 引用 引用 | ジャーナル: Science / 年: 2016 タイトル: Atomic structure of Hsp90-Cdc37-Cdk4 reveals that Hsp90 traps and stabilizes an unfolded kinase. 著者: Kliment A Verba / Ray Yu-Ruei Wang / Akihiko Arakawa / Yanxin Liu / Mikako Shirouzu / Shigeyuki Yokoyama / David A Agard /   要旨: The Hsp90 molecular chaperone and its Cdc37 cochaperone help stabilize and activate more than half of the human kinome. However, both the mechanism by which these chaperones assist their "client" ...The Hsp90 molecular chaperone and its Cdc37 cochaperone help stabilize and activate more than half of the human kinome. However, both the mechanism by which these chaperones assist their "client" kinases and the reason why some kinases are addicted to Hsp90 while closely related family members are independent are unknown. Our structural understanding of these interactions is lacking, as no full-length structures of human Hsp90, Cdc37, or either of these proteins with a kinase have been elucidated. Here we report a 3.9 angstrom cryo-electron microscopy structure of the Hsp90-Cdc37-Cdk4 kinase complex. Surprisingly, the two lobes of Cdk4 are completely separated with the β4-β5 sheet unfolded. Cdc37 mimics part of the kinase N lobe, stabilizing an open kinase conformation by wedging itself between the two lobes. Finally, Hsp90 clamps around the unfolded kinase β5 strand and interacts with exposed N- and C-lobe interfaces, protecting the kinase in a trapped unfolded state. On the basis of this structure and an extensive amount of previously collected data, we propose unifying conceptual and mechanistic models of chaperone-kinase interactions. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_3339.map.gz | 57 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-3339-v30.xmlemd-3339.xml | 15.6 KB 15.6 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_3339_fsc.xml | 10.7 KB | 表示 | FSCデータファイル |
| 画像 | emd_3339.tif | 144.2 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-3339ftp://ftp.pdbj.org/pub/emdb/structures/EMD-3339 http://ftp.pdbj.org/pub/emdb/structures/EMD-3339ftp://ftp.pdbj.org/pub/emdb/structures/EMD-3339 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_3339_validation.pdf.gz | 250.6 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_3339_full_validation.pdf.gz | 249.8 KB | 表示 | |
| XML形式データ | emd_3339_validation.xml.gz | 10.7 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-3339ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-3339 | HTTPS FTP |
-関連構造データ







-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |




















-マップ
| ファイル | ダウンロード / ファイル: emd_3339.map.gz / 形式: CCP4 / 大きさ: 62.5 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Reconstruction of Hsp90:Cdc37:Cdk4 complex. Part of series of maps, the highest resolution map being EMD-3337. This one is same as EMD-3337, but un-sharpened and filtered to 5.5A. This map has more coherent density for Cdk4 C-lobe. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.315 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : Complex of Human Hsp90 beta, human Cdc37 and human Cdk4
| 全体 | 名称: Complex of Human Hsp90 beta, human Cdc37 and human Cdk4 |
|---|---|
| 要素 |
|
-超分子 #1000: Complex of Human Hsp90 beta, human Cdc37 and human Cdk4
| 超分子 | 名称: Complex of Human Hsp90 beta, human Cdc37 and human Cdk4 タイプ: sample / ID: 1000 / 詳細: All three proteins were co-expressed in Sf9 cells. 集合状態: One Hsp90 homodimer binds to one Cdc37 and one Cdk4 Number unique components: 3 |
|---|---|
| 分子量 | 実験値: 245 KDa / 理論値: 245 KDa / 手法: As cloned, verified by SDS-PAGE |
-分子 #1: Heat Shock Protein HSP 90 beta
| 分子 | 名称: Heat Shock Protein HSP 90 beta / タイプ: protein_or_peptide / ID: 1 / Name.synonym: Hsp90 / コピー数: 2 / 集合状態: Dimer / 組換発現: Yes |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) / 別称: Human / 細胞中の位置: cytoplasm |
| 分子量 | 理論値: 83 KDa |
| 組換発現 | 生物種:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)組換プラスミド: pFastBacHT |
| 配列 | UniProtKB: Heat shock protein HSP 90-beta / GO: citrulline metabolic process / InterPro: Heat shock protein Hsp90 family |
-分子 #2: Hsp90 co-chaperone Cdc37
| 分子 | 名称: Hsp90 co-chaperone Cdc37 / タイプ: protein_or_peptide / ID: 2 / Name.synonym: Cdc37 / コピー数: 1 / 組換発現: Yes |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) / 別称: Human / 細胞中の位置: throughout |
| 分子量 | 理論値: 44.5 KDa |
| 組換発現 | 生物種: Spodoptera frugiperda (ツマジロクサヨトウ) 組換プラスミド: pFastBacHT |
| 配列 | UniProtKB: Hsp90 co-chaperone Cdc37 / GO: GO: 0000002 |
-分子 #3: Cyclin-dependent kinase 4
| 分子 | 名称: Cyclin-dependent kinase 4 / タイプ: protein_or_peptide / ID: 3 / Name.synonym: Cdk4 / コピー数: 1 / 組換発現: Yes |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) / 別称: Human / 細胞中の位置: throughout |
| 分子量 | 理論値: 33.7 KDa |
| 組換発現 | 生物種: Spodoptera frugiperda (ツマジロクサヨトウ) 組換プラスミド: pFastBacHT |
| 配列 | UniProtKB: Cyclin-dependent kinase 4 / GO: very long-chain fatty acid metabolic process / InterPro: Protein kinase domain |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 0.27 mg/mL |
|---|---|
| 緩衝液 | pH: 7.5 詳細: 20mM Tris-HCl (pH 7.5), 150 mM NaCl, 10 mM KCl, 10 mM MgCl2, 20 mM Na2MoO4, 2mM DTT, 0.085mM DDM |
| グリッド | 詳細: Glow discharged for 30 sec, C-flat 400 mesh 1.2/1.3 thick carbon grids (Protochips) |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 90 % / チャンバー内温度: 95 K / 装置: FEI VITROBOT MARK III / 手法: Single blot from 4 to 6 seconds, at 20C |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| アライメント法 | Legacy - 非点収差: At high mag via FT. |
| 日付 | 2014年11月25日 |
| 撮影 | カテゴリ: CCD フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 実像数: 3718 / 平均電子線量: 44 e/Å2 / 詳細: 38 frames, 7.6 seconds total exposure / ビット/ピクセル: 8 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: OTHER / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 3.8 µm / 最小 デフォーカス(公称値): 1.4 µm / 倍率(公称値): 22500 |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 詳細 | Image stacks were corrected for motion and summed as described previously, resulting in binned sums (1.315A/pix). For particle picking the images were binned to 5.2A/pix and Gaussian bandpass filtered between 15A and 500A using EMAN2. SamViewer template based picking was then used to pick particles from all the micrographs, followed by manual review of all the picks. After such procedure 802877 particles were picked in total and extracted from images binned to 2.6A/pix. CTFFIND4 was used to estimate defocus parameters for all the images. Relion 1.4 was used for all the following steps unless noted otherwise. Reference free 2D classification into 300 classes for 75 iterations was performed followed by manual examination of the resulting class averages. Low resolution/signal to noise/feature class averages and contributing particles were discarded, resulting in 670000 particles left. The resulting particles were 3D classified into 4 classes resulting in two classes having high-resolution features (390000 particles). At this stage particles were extracted from 1.315A/pix micrographs and all the following processing was done with these particles. Using 3D Auto-refine in Relion 1.4, a reconstruction was obtained from 390000 particles resulting from 3D classification above (using highest resolution 3D class as initial model, low pass filtered to 20A). Using the resulting parameters, the particles were further drift corrected per particle and dose weighted using the Particle Polishing feature. The B-factor weighing curve was fit by a polynomial (with a rationale that such a curve should be smooth) and used to generate new weighting parameters for Particle Polishing, with which 390000 particles were then polished. All further data processing was done using the polished particles. Re-refinement of the 390000 particles after polishing yielded the map at about 4A resolution (determined using gold standard FSC in the PostProcessing tab). Raw particles were sharpened with a B-factor of -50, low pass filtered with Gaussian filter to 3A and the refinement was continued for 10 more iterations (until convergence) with these particles (the rationale was that due to extremely low noise levels of K2 direct detector, this would yield more accurate alignments due to presence of more high resolution data in the images). This reconstruction was not sharpened and was filtered based on the spherical mask FSC resolution, 5.6A. Otherwise it is the same as EMD-3338. |
|---|---|
| 最終 再構成 | 想定した対称性 - 点群: C1 (非対称) / 解像度のタイプ: BY AUTHOR / 解像度: 5.6 Å / 解像度の算出法: OTHER / ソフトウェア - 名称: Relion / 使用した粒子像数: 388688 |
| 最終 2次元分類 | クラス数: 1 |
| FSC曲線 (解像度の算出) |  |
-原子モデル構築 1
| 初期モデル | PDB ID: Chain - #0 - Chain ID: A / Chain - #1 - Chain ID: B / Chain - #2 - Chain ID: E / Chain - #3 - Chain ID: K |
|---|---|
| ソフトウェア | 名称: Chimera |
| 詳細 | The model was built using Rosetta into EMD-3338 and its sister EMD-3337 (depending on the region), but was refined into the EMD-3337 map only. Then the resulting model was rigid body fit into this map in Chimera. |
| 精密化 | 空間: REAL / プロトコル: RIGID BODY FIT |
