 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5fwp | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Atomic cryoEM structure of Hsp90-Cdc37-Cdk4 complex | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | CHAPERONE / HSP90 / CDC37 / CDK4 / KINASE / UNFOLDING | |||||||||
| Function / homology |  Function and homology information Function and homology informationcyclin D3-CDK4 complex / cyclin D1-CDK4 complex / cyclin D2-CDK4 complex / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Drug-mediated inhibition of CDK4/CDK6 activity / regulation of type II interferon-mediated signaling pathway / regulation of transcription initiation by RNA polymerase II ...cyclin D3-CDK4 complex / cyclin D1-CDK4 complex / cyclin D2-CDK4 complex / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 / Evasion of Oncogene Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Evasion of Oxidative Stress Induced Senescence Due to Defective p16INK4A binding to CDK4 and CDK6 / Drug-mediated inhibition of CDK4/CDK6 activity / regulation of type II interferon-mediated signaling pathway / regulation of transcription initiation by RNA polymerase II / regulation of type B pancreatic cell proliferation / HSP90-CDC37 chaperone complex / negative regulation of proteasomal protein catabolic process / Aryl hydrocarbon receptor signalling / aryl hydrocarbon receptor complex / cellular response to ionomycin / histone methyltransferase binding / Transcriptional regulation by RUNX2 / dynein axonemal particle / cellular response to phorbol 13-acetate 12-myristate / positive regulation of type 2 mitophagy / receptor ligand inhibitor activity / protein kinase regulator activity / positive regulation of protein localization to cell surface / regulation of cyclin-dependent protein serine/threonine kinase activity / ATP-dependent protein binding / cyclin-dependent protein serine/threonine kinase regulator activity / post-transcriptional regulation of gene expression / Respiratory syncytial virus genome replication / telomerase holoenzyme complex assembly / Drug-mediated inhibition of ERBB2 signaling / Resistance of ERBB2 KD mutants to trastuzumab / Resistance of ERBB2 KD mutants to sapitinib / Resistance of ERBB2 KD mutants to tesevatinib / Resistance of ERBB2 KD mutants to neratinib / Resistance of ERBB2 KD mutants to osimertinib / Resistance of ERBB2 KD mutants to afatinib / Resistance of ERBB2 KD mutants to AEE788 / Resistance of ERBB2 KD mutants to lapatinib / Drug resistance in ERBB2 TMD/JMD mutants / Uptake and function of diphtheria toxin / PTK6 Regulates Cell Cycle / regulation of type I interferon-mediated signaling pathway / positive regulation of transforming growth factor beta receptor signaling pathway / dendritic growth cone / TPR domain binding / Defective binding of RB1 mutants to E2F1,(E2F2, E2F3) / Assembly and release of respiratory syncytial virus (RSV) virions / Sema3A PAK dependent Axon repulsion / The NLRP3 inflammasome / regulation of protein ubiquitination / protein phosphatase activator activity / HSF1-dependent transactivation / protein folding chaperone complex / response to unfolded protein / bicellular tight junction / positive regulation of G2/M transition of mitotic cell cycle / cyclin-dependent kinase / Attenuation phase / cyclin-dependent protein serine/threonine kinase activity / HSF1 activation / chaperone-mediated protein complex assembly / axonal growth cone / RHOBTB2 GTPase cycle / telomere maintenance via telomerase / Purinergic signaling in leishmaniasis infection / cyclin-dependent protein kinase holoenzyme complex / protein targeting / regulation of G2/M transition of mitotic cell cycle / Signaling by ERBB2 / supramolecular fiber organization / heat shock protein binding / peptide binding / DNA polymerase binding / protein folding chaperone / cellular response to interleukin-4 / ESR-mediated signaling / negative regulation of proteasomal ubiquitin-dependent protein catabolic process / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / placenta development / Constitutive Signaling by Overexpressed ERBB2 / cyclin binding / nitric-oxide synthase regulator activity / positive regulation of cell differentiation / ATP-dependent protein folding chaperone / Ubiquitin-dependent degradation of Cyclin D / Signaling by ERBB2 TMD/JMD mutants / G1/S transition of mitotic cell cycle / Constitutive Signaling by EGFRvIII / Hsp90 protein binding / Signaling by ERBB2 ECD mutants / Transcriptional regulation of white adipocyte differentiation / Signaling by ERBB2 KD Mutants / Regulation of actin dynamics for phagocytic cup formation / DDX58/IFIH1-mediated induction of interferon-alpha/beta / Oncogene Induced Senescence / Meiotic recombination / Regulation of necroptotic cell death / tau protein binding / positive regulation of fibroblast proliferation Similarity search - Function | |||||||||
| Biological species |  HOMO SAPIENS (human) HOMO SAPIENS (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 7.2 Å | |||||||||
 Authors Authors | Verba, K.A. / Wang, R.Y.R. / Arakawa, A. / Liu, Y. / Yokoyama, S. / Agard, D.A. | |||||||||
 Citation Citation | Journal: Science / Year: 2016 Title: Atomic structure of Hsp90-Cdc37-Cdk4 reveals that Hsp90 traps and stabilizes an unfolded kinase. Authors: Kliment A Verba / Ray Yu-Ruei Wang / Akihiko Arakawa / Yanxin Liu / Mikako Shirouzu / Shigeyuki Yokoyama / David A Agard /   Abstract: The Hsp90 molecular chaperone and its Cdc37 cochaperone help stabilize and activate more than half of the human kinome. However, both the mechanism by which these chaperones assist their "client" ...The Hsp90 molecular chaperone and its Cdc37 cochaperone help stabilize and activate more than half of the human kinome. However, both the mechanism by which these chaperones assist their "client" kinases and the reason why some kinases are addicted to Hsp90 while closely related family members are independent are unknown. Our structural understanding of these interactions is lacking, as no full-length structures of human Hsp90, Cdc37, or either of these proteins with a kinase have been elucidated. Here we report a 3.9 angstrom cryo-electron microscopy structure of the Hsp90-Cdc37-Cdk4 kinase complex. Surprisingly, the two lobes of Cdk4 are completely separated with the β4-β5 sheet unfolded. Cdc37 mimics part of the kinase N lobe, stabilizing an open kinase conformation by wedging itself between the two lobes. Finally, Hsp90 clamps around the unfolded kinase β5 strand and interacts with exposed N- and C-lobe interfaces, protecting the kinase in a trapped unfolded state. On the basis of this structure and an extensive amount of previously collected data, we propose unifying conceptual and mechanistic models of chaperone-kinase interactions. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5fwp.cif.gz | 593.2 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5fwp.ent.gz | 475.6 KB | Display | PDB format |
| PDBx/mmJSON format | 5fwp.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/fw/5fwpftp://data.pdbj.org/pub/pdb/validation_reports/fw/5fwp | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  3340MC  3337C  3338C  3339C  3341C  3342C  3343C  3344C  5fwkC  5fwlC  5fwmC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj









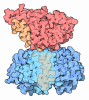


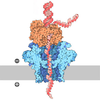









- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 83645.539 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Details: ATP / Source: (gene. exp.) HOMO SAPIENS (human) / Plasmid: PFASTBACHT / Production host:   SPODOPTERA FRUGIPERDA (fall armyworm) / References: UniProt: P08238 SPODOPTERA FRUGIPERDA (fall armyworm) / References: UniProt: P08238#2: Protein | | Mass: 44622.363 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) HOMO SAPIENS (human) / Plasmid: PFASTBACHT / Production host: SPODOPTERA FRUGIPERDA (fall armyworm) / References: UniProt: Q16543#3: Protein | | Mass: 34520.629 Da / Num. of mol.: 1 / Fragment: KINASE DOMAIN Source method: isolated from a genetically manipulated source Source: (gene. exp.) HOMO SAPIENS (human) / Plasmid: PFASTBACHT / Production host: SPODOPTERA FRUGIPERDA (fall armyworm) / References: UniProt: P11802#4: Chemical |   Mass: 507.181 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM#5: Chemical |   Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: MgHas protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.245 MDa / Experimental value: NO | ||||||||||||||||||||
| Buffer solution | pH: 7.5 Details: 20mM Tris-HCl (pH 7.5), 150 mM NaCl, 10 mM KCl, 10 mM MgCl2, 20 mM Na2MoO4, 2mM DTT, 0.085mM DDM | ||||||||||||||||||||
| Specimen | Conc.: 0.27 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||
| Specimen support | Details: HOLEY CARBON | ||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK III / Cryogen name: ETHANE / Temp: 95 K / Humidity: 90 % / Chamber temperature: 293 K / Details: LIQUID ETHANE / Method: Single blot from 4 to 6 seconds, at 20C |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS / Date: Nov 25, 2014 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 22500 X / Nominal defocus max: 3800 nm / Nominal defocus min: 1400 nm / Cs: 2.7 mm |
| Specimen holder | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 7.6 sec. / Electron dose: 44 e/Å2 / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of real images: 3718 / Details: 38 frames |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Image processing | Details: Image stacks were corrected for motion and summed as described previously, resulting in binned sums (1.315A/pix). For particle picking the images were binned to 5.2A/pix and Gaussian ...Details: Image stacks were corrected for motion and summed as described previously, resulting in binned sums (1.315A/pix). For particle picking the images were binned to 5.2A/pix and Gaussian bandpass filtered between 15A and 500A using EMAN2. SamViewer template based picking was then used to pick particles from all the micrographs, followed by manual review of all the picks. After such procedure 802877 particles were picked in total and extracted from images binned to 2.6A/pix. CTFFIND4 was used to estimate defocus parameters for all the images. Relion 1.4 was used for all the following steps unless noted otherwise. Reference free 2D classification into 300 classes for 75 iterations was performed followed by manual examination of the resulting class averages. Low resolution/signal to noise/feature class averages and contributing particles were discarded, resulting in 670000 particles left. The resulting particles were 3D classified into 4 classes resulting in two classes having high-resolution features (390000 particles). At this stage particles were extracted from 1.315A/pix micrographs and all the following processing was done with these particles. Using 3D Auto-refine in Relion 1.4, a reconstruction was obtained from 390000 particles resulting from 3D classification above (using highest resolution 3D class as initial model, low pass filtered to 20A). Using the resulting parameters, the particles were further drift corrected per particle and dose weighted using the Particle Polishing feature. The B-factor weighing curve was fit by a polynomial (with a rationale that such a curve should be smooth) and used to generate new weighting parameters for Particle Polishing, with which 390000 particles were then polished. All further data processing was done using the polished particles. Re-refinement of the 390000 particles after polishing yielded the map at about 4A resolution (determined using gold standard FSC in the PostProcessing tab). The 390000 particles were then 3D classified into four different classes without particle re-alignment, using the alignment parameters from 4A reconstruction. Particles contributing to each of the four classes were grouped and a full 3D refinement with a spherical 200A mask was performed with each of the four groups of particles using the same initial model, low pass filtered to 20A. This is one of the resulting reconstructions. | ||||||||||||
| Particle selection | Num. of particles selected: 114683 | ||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||
| 3D reconstruction | Method: FSC 0.143 CUT-OFF / Resolution: 7.2 Å / Num. of particles: 114683 / Actual pixel size: 1.315 Å Details: SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-3340. (DEPOSITION ID: 14281). Refinement type: HALF-MAPS REFINED INDEPENDENTLY / Symmetry type: POINT | ||||||||||||
| Atomic model building | B value: 141 / Protocol: FLEXIBLE FIT / Space: REAL Details: Cdc37 crystal structure from PDB:1US7 was fit into EMD-3340 manually and then with UCSF Chimera "Fit In Map" tool. The model was truncated at residue 260, as there was no reliable density ...Details: Cdc37 crystal structure from PDB:1US7 was fit into EMD-3340 manually and then with UCSF Chimera "Fit In Map" tool. The model was truncated at residue 260, as there was no reliable density for the rest of the crystal structure. PDB:5FWK and the above fit crystal structure for residues 148-260 were loaded in Coot. The residues 133-147 were built in by hand into the Cdc37 Reconstruction and residues 245-260 (helix) were rotated as a rigid body. To relieve atomic clashes or bond length/angle distortions at the linker regions, the resulting model was subjected to "Cartesian space relax" protocol within EMD-3340 map using Rosetta. Final model was selected using the combined score of Rosetta all-atom physically-realistic score and electron density score. | ||||||||||||
| Atomic model building |
| ||||||||||||
| Refinement | Highest resolution: 7.2 Å | ||||||||||||
| Refinement step | Cycle: LAST / Highest resolution: 7.2 Å
|
