response to oxygen-glucose deprivation / hippocampal neuron apoptotic process / poly-ADP-D-ribose binding / positive regulation of cell growth involved in cardiac muscle cell development / NAD+-protein-serine ADP-ribosyltransferase activity / NAD DNA ADP-ribosyltransferase activity / DNA ADP-ribosylation / poly-ADP-D-ribose modification-dependent protein binding / HDR through MMEJ (alt-NHEJ) / NAD+ ADP-ribosyltransferase ...response to oxygen-glucose deprivation / hippocampal neuron apoptotic process / poly-ADP-D-ribose binding / positive regulation of cell growth involved in cardiac muscle cell development / NAD+-protein-serine ADP-ribosyltransferase activity / NAD DNA ADP-ribosyltransferase activity / DNA ADP-ribosylation / poly-ADP-D-ribose modification-dependent protein binding / HDR through MMEJ (alt-NHEJ) / NAD+ ADP-ribosyltransferase / protein auto-ADP-ribosylation / NAD+-protein-aspartate ADP-ribosyltransferase activity / protein poly-ADP-ribosylation / NAD+-protein-glutamate ADP-ribosyltransferase activity / DNA repair-dependent chromatin remodeling / NAD+-protein mono-ADP-ribosyltransferase activity / decidualization / site of DNA damage / NAD+ poly-ADP-ribosyltransferase activity / Transferases; Glycosyltransferases; Pentosyltransferases / POLB-Dependent Long Patch Base Excision Repair / negative regulation of tumor necrosis factor-mediated signaling pathway / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / nucleosome binding / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / extrinsic apoptotic signaling pathway / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / nucleotidyltransferase activity / telomere organization / Interleukin-7 signaling / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / DNA methylation / epigenetic regulation of gene expression / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / innate immune response in mucosa / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / Nonhomologous End-Joining (NHEJ) / lipopolysaccharide binding / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / HDMs demethylate histones / G2/M DNA damage checkpoint / base-excision repair / DNA Damage Recognition in GG-NER / NoRC negatively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / B-WICH complex positively regulates rRNA expression / PKMTs methylate histone lysines / Dual Incision in GG-NER / Meiotic recombination / Pre-NOTCH Transcription and Translation / Metalloprotease DUBs / Formation of Incision Complex in GG-NER / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / HCMV Early Events / antimicrobial humoral immune response mediated by antimicrobial peptide / structural constituent of chromatin / UCH proteinases / antibacterial humoral response / nucleosome / heterochromatin formation / double-strand break repair / nucleosome assembly / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / HATs acetylate histones / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Factors involved in megakaryocyte development and platelet production / chromatin organization / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / Oxidative Stress Induced Senescence / defense response to Gram-negative bacterium Similarity search - Function
National Institutes of Health/National Cancer Institute (NIH/NCI)
R01 CA218255
United States
Citation
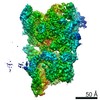
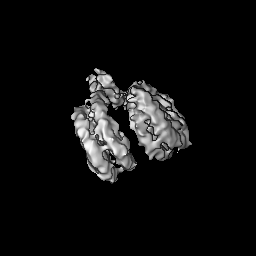
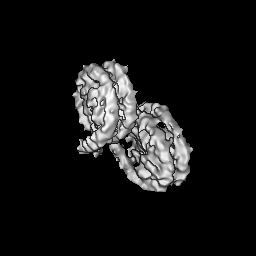
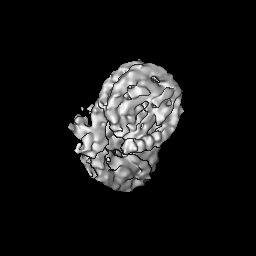
Journal: PLoS One / Year: 2020 Title: Bridging of nucleosome-proximal DNA double-strand breaks by PARP2 enhances its interaction with HPF1. Authors: Guillaume Gaullier / Genevieve Roberts / Uma M Muthurajan / Samuel Bowerman / Johannes Rudolph / Jyothi Mahadevan / Asmita Jha / Purushka S Rae / Karolin Luger / Abstract: Poly(ADP-ribose) Polymerase 2 (PARP2) is one of three DNA-dependent PARPs involved in the detection of DNA damage. Upon binding to DNA double-strand breaks, PARP2 uses nicotinamide adenine ...Poly(ADP-ribose) Polymerase 2 (PARP2) is one of three DNA-dependent PARPs involved in the detection of DNA damage. Upon binding to DNA double-strand breaks, PARP2 uses nicotinamide adenine dinucleotide to synthesize poly(ADP-ribose) (PAR) onto itself and other proteins, including histones. PAR chains in turn promote the DNA damage response by recruiting downstream repair factors. These early steps of DNA damage signaling are relevant for understanding how genome integrity is maintained and how their failure leads to genome instability or cancer. There is no structural information on DNA double-strand break detection in the context of chromatin. Here we present a cryo-EM structure of two nucleosomes bridged by human PARP2 and confirm that PARP2 bridges DNA ends in the context of nucleosomes bearing short linker DNA. We demonstrate that the conformation of PARP2 bound to damaged chromatin provides a binding platform for the regulatory protein Histone PARylation Factor 1 (HPF1), and that the resulting HPF1•PARP2•nucleosome complex is enzymatically active. Our results contribute to a structural view of the early steps of the DNA damage response in chromatin.
History
Deposition
Oct 27, 2019
-
Header (metadata) release
Nov 6, 2019
-
Map release
Oct 14, 2020
-
Update
Mar 6, 2024
-
Current status
Mar 6, 2024
Processing site: RCSB / Status: Released
-
Structure visualization
Movie
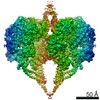
Surface view with section colored by density value
EMPIAR-10336 (Title: Structure of two nucleosomes bridged by human PARP2 / Data size: 677.3 Data #1: Single-particle images of the PARP2-nucleosome complex [picked particles - single frame - unprocessed] Data #2: Unaligned multi-frame micrographs of the PARP2-nucleosome complex [micrographs - multiframe])
Protein or peptide: Poly [ADP-ribose] polymerase 2
+
Supramolecule #1: Two nucleosomes bridged by PARP2
Supramolecule
Name: Two nucleosomes bridged by PARP2 / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all Details: Two nucleosomes bridged by human PARP2. Full-length PARP2 was used to prepare the complex, but only the WGR domain showed density in the map.
Name: Widom 601 DNA (160-MER) / type: dna / ID: 1 Details: Widom 601 nucleosome positioning sequence with 7 and 11 additional terminal base pairs Number of copies: 2 / Classification: DNA
Details: Buffer was pH-adjusted and filtered through a 0.22 um filter.
Grid
Model: Quantifoil R1.2/1.3 / Material: COPPER / Mesh: 300 / Support film - Material: CARBON / Support film - topology: HOLEY ARRAY / Pretreatment - Type: GLOW DISCHARGE / Pretreatment - Time: 30 sec. / Pretreatment - Atmosphere: AIR
Vitrification
Cryogen name: ETHANE / Chamber humidity: 95 % / Chamber temperature: 277 K / Instrument: FEI VITROBOT MARK IV / Details: Blot time 2s, blot force 0.
-
Electron microscopy
Microscope
FEI TECNAI F30
Image recording
Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Detector mode: COUNTING / Digitization - Frames/image: 1-40 / Number grids imaged: 1 / Number real images: 632 / Average exposure time: 4.0 sec. / Average electron dose: 56.1 e/Å2
Electron beam
Acceleration voltage: 300 kV / Electron source: FIELD EMISSION GUN
Electron optics
Illumination mode: OTHER / Imaging mode: BRIGHT FIELD
Sample stage
Specimen holder model: GATAN 626 SINGLE TILT LIQUID NITROGEN CRYO TRANSFER HOLDER Cooling holder cryogen: NITROGEN
Experimental equipment
Model: Tecnai F30 / Image courtesy: FEI Company
+
Image processing
Particle selection
Number selected: 226250 Details: Picked using crYOLO version 1.3.1 with a model trained on the same dataset (with the generic model provided with crYOLO used as pre-trained weights).
Startup model
Type of model: INSILICO MODEL In silico model: Model generated with the InitialModel procedure in RELION
Final reconstruction
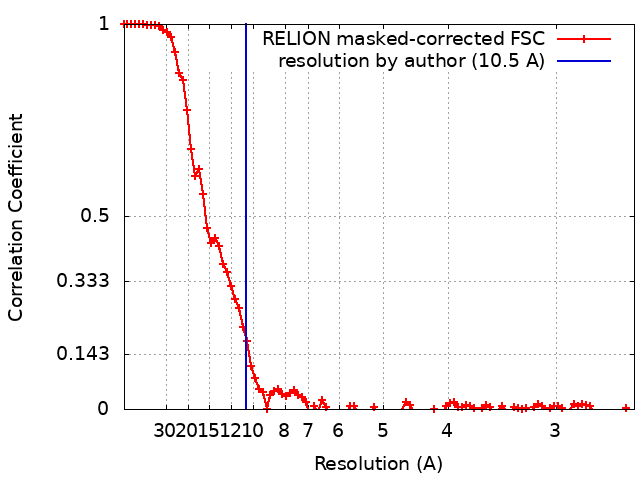
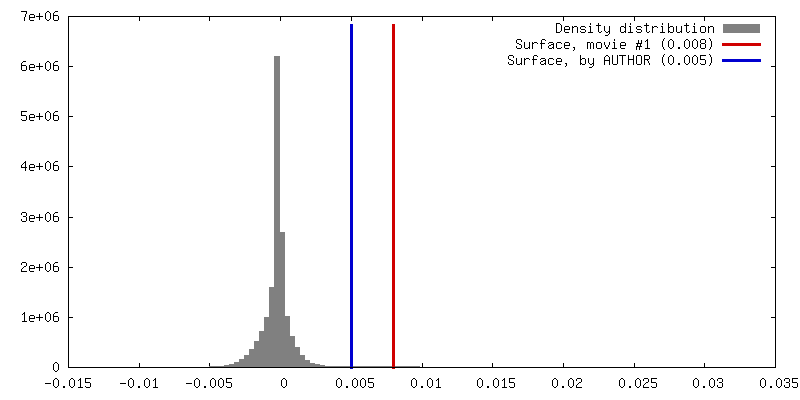
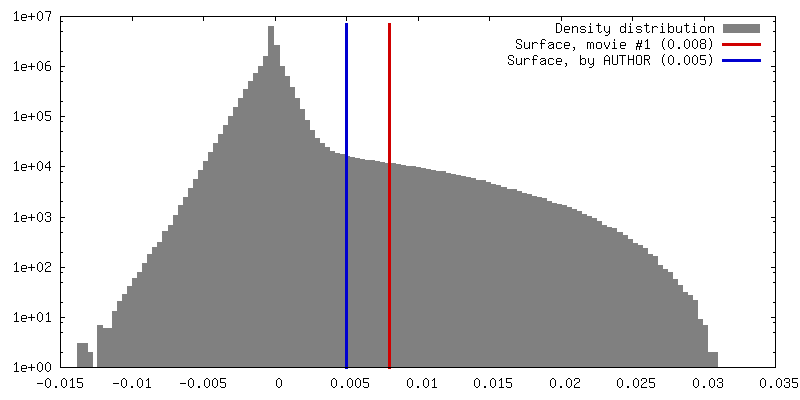
Number classes used: 1 / Applied symmetry - Point group: C1 (asymmetric) / Resolution.type: BY AUTHOR / Resolution: 10.5 Å / Resolution method: FSC 0.143 CUT-OFF / Software - Name: RELION (ver. 3.0) / Number images used: 27889
Initial angle assignment
Type: MAXIMUM LIKELIHOOD / Software - Name: RELION (ver. 3.0)
Final angle assignment
Type: MAXIMUM LIKELIHOOD / Software - Name: RELION (ver. 3.0)
Final 3D classification
Number classes: 5 / Avg.num./class: 33753 / Software - Name: RELION (ver. 3.0) / Details: 27889 particles were selected for refinement.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Map data
Map data Sample
Sample Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human) / synthetic construct (others)
Homo sapiens (human) / synthetic construct (others) Authors
Authors United States, 1 items
United States, 1 items  Citation
Citation Structure visualization
Structure visualization
 Downloads & links
Downloads & links emd_20864.png
emd_20864.png http://ftp.pdbj.org/pub/emdb/structures/EMD-20864
http://ftp.pdbj.org/pub/emdb/structures/EMD-20864

















 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)



























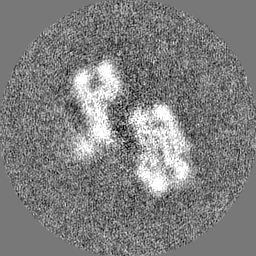






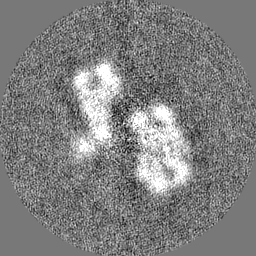



 Sample components
Sample components
 Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN