 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6z6k | ||||||
|---|---|---|---|---|---|---|---|
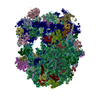
| タイトル | Cryo-EM structure of yeast reconstituted Lso2 bound to 80S ribosomes | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / hibernation | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報cytoplasmic translational elongation / ribosome hibernation / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / cellular response to nutrient / Negative regulators of DDX58/IFIH1 signaling / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / positive regulation of translational fidelity / RMTs methylate histone arginines / Protein methylation ...cytoplasmic translational elongation / ribosome hibernation / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / cellular response to nutrient / Negative regulators of DDX58/IFIH1 signaling / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / positive regulation of translational fidelity / RMTs methylate histone arginines / Protein methylation / mTORC1-mediated signalling / Protein hydroxylation / ribosome-associated ubiquitin-dependent protein catabolic process / GDP-dissociation inhibitor activity / positive regulation of nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay / pre-mRNA 5'-splice site binding / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / Ribosomal scanning and start codon recognition / preribosome, small subunit precursor / nonfunctional rRNA decay / response to cycloheximide / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / Major pathway of rRNA processing in the nucleolus and cytosol / mRNA destabilization / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / negative regulation of translational frameshifting / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / negative regulation of mRNA splicing, via spliceosome / Formation of a pool of free 40S subunits / preribosome, large subunit precursor / positive regulation of protein kinase activity / L13a-mediated translational silencing of Ceruloplasmin expression / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translational elongation / ribosomal large subunit export from nucleus / 90S preribosome / G-protein alpha-subunit binding / Ub-specific processing proteases / ribosomal subunit export from nucleus / regulation of translational fidelity / protein-RNA complex assembly / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translational termination / maturation of LSU-rRNA / ribosomal small subunit export from nucleus / translation regulator activity / translation repressor activity / DNA-(apurinic or apyrimidinic site) endonuclease activity / rescue of stalled ribosome / cellular response to amino acid starvation / protein kinase C binding / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosome assembly / ribosomal large subunit biogenesis / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / translational initiation / small-subunit processome / macroautophagy / maintenance of translational fidelity / modification-dependent protein catabolic process / protein tag activity / cytoplasmic stress granule / rRNA processing / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / 5S rRNA binding / ribosomal large subunit assembly / cytosolic small ribosomal subunit / large ribosomal subunit rRNA binding / small ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / intracellular iron ion homeostasis / cytoplasmic translation / negative regulation of translation / rRNA binding / protein ubiquitination / structural constituent of ribosome / ribosome / G protein-coupled receptor signaling pathway / translation / negative regulation of gene expression / response to antibiotic / mRNA binding / ubiquitin protein ligase binding / nucleolus / mitochondrion / RNA binding / zinc ion binding / nucleoplasm / nucleus / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.4 Å | ||||||
 データ登録者 データ登録者 | Wells, J.N. / Buschauer, R. / Mackens-Kiani, T. / Best, K. / Kratzat, H. / Berninghausen, O. / Becker, T. / Cheng, J. / Beckmann, R. | ||||||
 引用 引用 | ジャーナル: PLoS Biol / 年: 2020 タイトル: Structure and function of yeast Lso2 and human CCDC124 bound to hibernating ribosomes. 著者: Jennifer N Wells / Robert Buschauer / Timur Mackens-Kiani / Katharina Best / Hanna Kratzat / Otto Berninghausen / Thomas Becker / Wendy Gilbert / Jingdong Cheng / Roland Beckmann /   要旨: Cells adjust to nutrient deprivation by reversible translational shutdown. This is accompanied by maintaining inactive ribosomes in a hibernation state, in which they are bound by proteins with ...Cells adjust to nutrient deprivation by reversible translational shutdown. This is accompanied by maintaining inactive ribosomes in a hibernation state, in which they are bound by proteins with inhibitory and protective functions. In eukaryotes, such a function was attributed to suppressor of target of Myb protein 1 (Stm1; SERPINE1 mRNA-binding protein 1 [SERBP1] in mammals), and recently, late-annotated short open reading frame 2 (Lso2; coiled-coil domain containing short open reading frame 124 [CCDC124] in mammals) was found to be involved in translational recovery after starvation from stationary phase. Here, we present cryo-electron microscopy (cryo-EM) structures of translationally inactive yeast and human ribosomes. We found Lso2/CCDC124 accumulating on idle ribosomes in the nonrotated state, in contrast to Stm1/SERBP1-bound ribosomes, which display a rotated state. Lso2/CCDC124 bridges the decoding sites of the small with the GTPase activating center (GAC) of the large subunit. This position allows accommodation of the duplication of multilocus region 34 protein (Dom34)-dependent ribosome recycling system, which splits Lso2-containing, but not Stm1-containing, ribosomes. We propose a model in which Lso2 facilitates rapid translation reactivation by stabilizing the recycling-competent state of inactive ribosomes. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6z6k.cif.gz | 4.3 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6z6k.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 6z6k.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6z6k_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6z6k_full_validation.pdf.gz | 1.5 MB | 表示 | |
| XML形式データ | 6z6k_validation.xml.gz | 317.7 KB | 表示 | |
| CIF形式データ | 6z6k_validation.cif.gz | 556.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/z6/6z6kftp://data.pdbj.org/pub/pdb/validation_reports/z6/6z6k | HTTPS FTP |
-関連構造データ


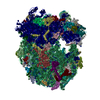
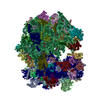
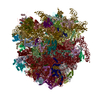
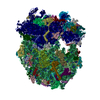




-リンク
 PDBj
PDBj































- 集合体
集合体
| 登録構造単位 | 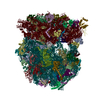
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 4種, 4分子 C2C1C4C3
| #1: RNA鎖 | 分子量: 579761.938 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: GenBank: 874346701 |
|---|---|
| #3: RNA鎖 | 分子量: 1097493.875 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: GenBank: 834774822 |
| #4: RNA鎖 | 分子量: 38951.105 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: GenBank: 1329886537 |
| #5: RNA鎖 | 分子量: 50682.922 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: GenBank: 1331532632 |
-タンパク質 , 5種, 5分子 C5SFSfSgLm
| #2: タンパク質 | 分子量: 10391.823 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: LSO2, YGR169C-A / 発現宿主:  |
|---|---|
| #11: タンパク質 | 分子量: 25072.600 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: UniProt: A0A1L4AA68 |
| #37: タンパク質 | 分子量: 17254.227 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: UniProt: P05759 |
| #38: タンパク質 | 分子量: 34841.219 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: UniProt: P38011 |
| #76: タンパク質 | 分子量: 14583.077 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 参照: UniProt: P0CH08 |
+40S ribosomal protein ... , 30種, 30分子 SASBSCSDSESGSHSISJSKSLSMSNSOSPSQSRSSSTSUSVSWSXSYSZSaSbScSdSe
+60S ribosomal protein ... , 40種, 40分子 LALBLCLDLELFLGLHLILJLLLMLNLOLPLQLRLSLTLULVLWLXLYLZLaLbLcLdLe...
-非ポリマー , 1種, 9分子 
| #80: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 9 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 9 / 由来タイプ: 合成 / 式: Zn |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) | 生物種: | ||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 | ||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 28 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 88523 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||
| 精密化 | 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 拘束条件 |
|
