 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
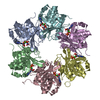
| 登録情報 | データベース: PDB / ID: 5muw | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Atomic structure of P4 packaging enzyme fitted into a localized reconstruction of bacteriophage phi6 vertex | ||||||
 要素 要素 | Packaging enzyme P4 | ||||||
 キーワード キーワード | HYDROLASE / packaging / ATPase / vertex / hyrdolase | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報viral procapsid / viral genome packaging / ribonucleoside triphosphate phosphatase activity / viral capsid / nucleoside-triphosphate phosphatase / ATP binding 類似検索 - 分子機能 | ||||||
| 生物種 |  Pseudomonas phage phi6 (ファージ) Pseudomonas phage phi6 (ファージ) | ||||||
| 手法 | 電子顕微鏡法 /  シンクロトロン / 単粒子再構成法 / 分子置換 / クライオ電子顕微鏡法 / 解像度: 9.1 Å シンクロトロン / 単粒子再構成法 / 分子置換 / クライオ電子顕微鏡法 / 解像度: 9.1 Å | ||||||
 データ登録者 データ登録者 | Sun, Z. / El Omari, K. / Sun, X. / Ilca, S. / Kotecha, A. / Stuart, D.I. / Poranen, M.M. / Huiskonen, J.T. | ||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2017 タイトル: Double-stranded RNA virus outer shell assembly by bona fide domain-swapping. 著者: Zhaoyang Sun / Kamel El Omari / Xiaoyu Sun / Serban L Ilca / Abhay Kotecha / David I Stuart / Minna M Poranen / Juha T Huiskonen /   要旨: Correct outer protein shell assembly is a prerequisite for virion infectivity in many multi-shelled dsRNA viruses. In the prototypic dsRNA bacteriophage φ6, the assembly reaction is promoted by ...Correct outer protein shell assembly is a prerequisite for virion infectivity in many multi-shelled dsRNA viruses. In the prototypic dsRNA bacteriophage φ6, the assembly reaction is promoted by calcium ions but its biomechanics remain poorly understood. Here, we describe the near-atomic resolution structure of the φ6 double-shelled particle. The outer T=13 shell protein P8 consists of two alpha-helical domains joined by a linker, which allows the trimer to adopt either a closed or an open conformation. The trimers in an open conformation swap domains with each other. Our observations allow us to propose a mechanistic model for calcium concentration regulated outer shell assembly. Furthermore, the structure provides a prime exemplar of bona fide domain-swapping. This leads us to extend the theory of domain-swapping from the level of monomeric subunits and multimers to closed spherical shells, and to hypothesize a mechanism by which closed protein shells may arise in evolution. #1: ジャーナル: Nucleic Acids Res / 年: 2013タイトル: Tracking in atomic detail the functional specializations in viral RecA helicases that occur during evolution. 著者: Kamel El Omari / Christoph Meier / Denis Kainov / Geoff Sutton / Jonathan M Grimes / Minna M Poranen / Dennis H Bamford / Roman Tuma / David I Stuart / Erika J Mancini / 要旨: Many complex viruses package their genomes into empty protein shells and bacteriophages of the Cystoviridae family provide some of the simplest models for this. The cystoviral hexameric NTPase, P4, ...Many complex viruses package their genomes into empty protein shells and bacteriophages of the Cystoviridae family provide some of the simplest models for this. The cystoviral hexameric NTPase, P4, uses chemical energy to translocate single-stranded RNA genomic precursors into the procapsid. We previously dissected the mechanism of RNA translocation for one such phage, 12, and have now investigated three further highly divergent, cystoviral P4 NTPases (from 6, 8 and 13). High-resolution crystal structures of the set of P4s allow a structure-based phylogenetic analysis, which reveals that these proteins form a distinct subfamily of the RecA-type ATPases. Although the proteins share a common catalytic core, they have different specificities and control mechanisms, which we map onto divergent N- and C-terminal domains. Thus, the RNA loading and tight coupling of NTPase activity with RNA translocation in 8 P4 is due to a remarkable C-terminal structure, which wraps right around the outside of the molecule to insert into the central hole where RNA binds to coupled L1 and L2 loops, whereas in 12 P4, a C-terminal residue, serine 282, forms a specific hydrogen bond to the N7 of purines ring to confer purine specificity for the 12 enzyme. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5muw.cif.gz | 631.9 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5muw.ent.gz | 515.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5muw.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5muw_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5muw_full_validation.pdf.gz | 1.3 MB | 表示 | |
| XML形式データ | 5muw_validation.xml.gz | 51 KB | 表示 | |
| CIF形式データ | 5muw_validation.cif.gz | 75.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/mu/5muwftp://data.pdbj.org/pub/pdb/validation_reports/mu/5muw | HTTPS FTP |
-関連構造データ








-リンク
 PDBj
PDBj- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 32678.740 Da / 分子数: 6 / 断片: RESIDUES 1-309 / 由来タイプ: 組換発現 由来: (組換発現) Pseudomonas phage phi6 (ファージ)遺伝子: P4 / 発現宿主:  参照: UniProt: P11125, nucleoside-triphosphate phosphatase #2: 化合物 | ChemComp-CA /   分子量: 40.078 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Ca 分子量: 40.078 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Ca#3: 化合物 | ChemComp-ADP /   分子量: 427.201 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Pseudomonas phage phi6 / タイプ: VIRUS / Entity ID: #1 / 由来: NATURAL |
|---|---|
| 由来(天然) | 生物種: Pseudomonas phage phi6 (ファージ) |
| ウイルスについての詳細 | 中空か: NO / エンベロープを持つか: YES / 単離: SPECIES / タイプ: VIRION |
| 天然宿主 | 生物種: Pseudomonas syringae |
| 緩衝液 | pH: 7.2 |
| 試料 | 濃度: 3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | グリッドの材料: COPPER / グリッドのタイプ: C-flat |
| 急速凍結 | 凍結剤: ETHANE |
| 結晶 | マシュー密度: 2.3 Å3/Da / 溶媒含有率: 46 % |
| 結晶化 | pH: 8 / 詳細: 6% PEG 4000 AND 90 MM SODIUM ACETATE PH 4.5 / PH範囲: 8 |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI POLARA 300 |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(補正後): 37037 X / Calibrated defocus min: 300 nm / 最大 デフォーカス(補正後): 3000 nm / Cs: 2 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN / 試料ホルダーモデル: OTHER / 最高温度: 120 K / 最低温度: 80 K |
| 撮影 | 平均露光時間: 0.2 sec. / 電子線照射量: 0.7 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 QUANTUM (4k x 4k) 実像数: 900 |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum LS / エネルギーフィルター 上限: 20 eV / エネルギーフィルター 下限: 0 eV |
| 画像スキャン | サンプリングサイズ: 5 µm / 横: 3710 / 縦: 3710 / 動画フレーム数/画像: 22 / 利用したフレーム数/画像: 1-22 |
| 回折 | 平均測定温度: 100 K |
| 放射光源 | 由来: シンクロトロン / サイト: SRS / ビームライン: PX14.2 / 波長: 0.87 |
| 検出器 | タイプ: ADSC QUANTUM 4r / 検出器: CCD |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: electron |
| 放射波長 | 波長: 0.87 Å / 相対比: 1 |
| 反射 | 解像度: 2.8→20 Å / Num. obs: 72831 / % possible obs: 97 % / Observed criterion σ(I): 1.5 / 冗長度: 3 % / Biso Wilson estimate: 51.06 Å2 / Rmerge(I) obs: 0.19 / Net I/σ(I): 8.3 |
| 反射 シェル | 解像度: 2.8→2.9 Å / 冗長度: 2.6 % / Rmerge(I) obs: 0.86 / Mean I/σ(I) obs: 1.5 / % possible all: 88.2 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 159492 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 9.1 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 56448 / アルゴリズム: FOURIER SPACE / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL / Target criteria: Cross-correlation coefficient | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 構造決定の手法: 分子置換 / 解像度: 9.1→9.1 Å / Cor.coef. Fo:Fc: 0.916 / Cor.coef. Fo:Fc free: 0.895 / 交差検証法: THROUGHOUT / σ(F): 0 / SU Rfree Blow DPI: 0.378
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 56.95 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.477 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.8→19.99 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | 解像度: 2.8→2.87 Å / Total num. of bins used: 20
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: ELECTRON MICROSCOPY
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
