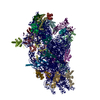
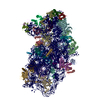
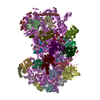
ジャーナル: Elife / 年: 2016 タイトル: Structural characterization of ribosome recruitment and translocation by type IV IRES. 著者: Jason Murray / Christos G Savva / Byung-Sik Shin / Thomas E Dever / V Ramakrishnan / Israel S Fernández / 要旨: Viral mRNA sequences with a type IV IRES are able to initiate translation without any host initiation factors. Initial recruitment of the small ribosomal subunit as well as two translocation steps ...Viral mRNA sequences with a type IV IRES are able to initiate translation without any host initiation factors. Initial recruitment of the small ribosomal subunit as well as two translocation steps before the first peptidyl transfer are essential for the initiation of translation by these mRNAs. Using electron cryomicroscopy (cryo-EM) we have structurally characterized at high resolution how the Cricket Paralysis Virus Internal Ribosomal Entry Site (CrPV-IRES) binds the small ribosomal subunit (40S) and the translocation intermediate stabilized by elongation factor 2 (eEF2). The CrPV-IRES restricts tvhe otherwise flexible 40S head to a conformation compatible with binding the large ribosomal subunit (60S). Once the 60S is recruited, the binary CrPV-IRES/80S complex oscillates between canonical and rotated states (Fernández et al., 2014; Koh et al., 2014), as seen for pre-translocation complexes with tRNAs. Elongation factor eEF2 with a GTP analog stabilizes the ribosome-IRES complex in a rotated state with an extra ~3 degrees of rotation. Key residues in domain IV of eEF2 interact with pseudoknot I (PKI) of the CrPV-IRES stabilizing it in a conformation reminiscent of a hybrid tRNA state. The structure explains how diphthamide, a eukaryotic and archaeal specific post-translational modification of a histidine residue of eEF2, is involved in translocation.
#89 - 2007年5月 アコニターゼと鉄調節タンパク質1 (Aconitase and Iron Regulatory Protein 1) 類似性 (1)
#241 - 2020年1月 20年の分子を振り返って (Twenty Years of Molecules) 類似性 (1)
-
集合体
登録構造単位
A: Ribosomal protein uS2 B: Ribosomal protein eS1 C: Ribosomal protein uS5 D: Ribosomal protein uS3 E: Ribosomal protein eS4 F: Ribosomal protein uS7 G: Ribosomal protein eS6 H: Ribosomal protein eS7 I: Ribosomal protein eS8 J: Ribosomal protein uS4 K: Ribosomal protein eS10 L: Ribosomal protein uS17 M: Ribosomal protein eS12 N: Ribosomal protein uS15 O: Ribosomal protein uS14 P: Ribosomal protein uS19 Q: Ribosomal protein uS9 R: Ribosomal protein eS17 S: Ribosomal protein uS13 T: Ribosomal protein eS19 U: Ribosomal protein uS10 V: Ribosomal protein eS21 W: Ribosomal protein uS8 X: Ribosomal protein uS21 Y: Ribosomal protein eS24 Z: Ribosomal protein eS25 a: Ribosomal protein eS26 b: Ribosomal protein eS27 c: Ribosomal protein eS28 d: Ribosomal protein eS29 e: Ribosomal protein eS30 f: Ribosomal protein eS31 g: Ribosomal protein RACK1 2: 18S ribosomal RNA i: Cricket paralysis virus IRES RNA ヘテロ分子
装置: FEI VITROBOT MARK II / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 298 K
-
電子顕微鏡撮影
実験機器
モデル: Titan Krios / 画像提供: FEI Company
顕微鏡
モデル: FEI TITAN KRIOS
電子銃
電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER
電子レンズ
モード: BRIGHT FIELD
試料ホルダ
試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER
撮影
電子線照射量: 25 e/Å2 / 検出モード: INTEGRATING フィルム・検出器のモデル: FEI FALCON II (4k x 4k)
-
解析
ソフトウェア
名称: REFMAC / バージョン: 5.8.0124 / 分類: 精密化
EMソフトウェア
ID
名称
バージョン
カテゴリ
1
EPU
粒子像選択
2
EPU
1.8
画像取得
4
CTFFIND
CTF補正
9
RELION
1.4
初期オイラー角割当
13
REFMAC
5.8
モデル精密化
CTF補正
タイプ: PHASE FLIPPING ONLY
3次元再構成
解像度: 3.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 54481 / アルゴリズム: BACK PROJECTION / 対称性のタイプ: POINT
原子モデル構築
プロトコル: OTHER / 空間: RECIPROCAL
精密化
解像度: 3.8→257.28 Å / Cor.coef. Fo:Fc: 0.848 / SU B: 43.619 / SU ML: 0.614 / σ(F): 0 / ESU R: 0.945 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS U VALUES : REFINED INDIVIDUALLY
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Cricket paralysis virus (ウイルス)
Cricket paralysis virus (ウイルス) Kluyveromyces lactis (酵母)
Kluyveromyces lactis (酵母) データ登録者
データ登録者 英国,
英国,  米国, 3件
米国, 3件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード



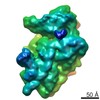

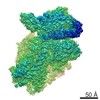
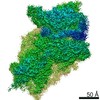

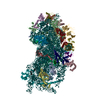
 PDBj
PDBj





























 集合体
集合体



 分子量: 24.305 Da / 分子数: 80 / 由来タイプ: 合成 / 式: Mg
分子量: 24.305 Da / 分子数: 80 / 由来タイプ: 合成 / 式: Mg 分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn
分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER
FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER 解析
解析