 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5it7: Structure of the Kluyveromyces lactis 80S ribosome in complex wit... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5it7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of the Kluyveromyces lactis 80S ribosome in complex with the cricket paralysis virus IRES and eEF2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Components Components |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Keywords Keywords | RIBOSOME / translocation / IRES / eEF2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationGDP-dissociation inhibitor activity / response to cycloheximide / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / preribosome, large subunit precursor / Formation of a pool of free 40S subunits / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / L13a-mediated translational silencing of Ceruloplasmin expression ...GDP-dissociation inhibitor activity / response to cycloheximide / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / preribosome, large subunit precursor / Formation of a pool of free 40S subunits / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / L13a-mediated translational silencing of Ceruloplasmin expression / translational elongation / ribosomal large subunit export from nucleus / 90S preribosome / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosomal subunit export from nucleus / maturation of LSU-rRNA / protein-RNA complex assembly / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translation regulator activity / cellular response to amino acid starvation / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / translational initiation / small-subunit processome / protein tag activity / rRNA processing / large ribosomal subunit / ribosomal small subunit assembly / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / 5S rRNA binding / ribosomal large subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / negative regulation of translation / rRNA binding / structural constituent of ribosome / protein ubiquitination / ribosome / translation / G protein-coupled receptor signaling pathway / ribonucleoprotein complex / response to antibiotic / mRNA binding / nucleolus / RNA binding / zinc ion binding / nucleus / cytoplasm / cytosol Similarity search - Function | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Biological species |  Cricket paralysis virus Cricket paralysis virus Kluyveromyces lactis (yeast) Kluyveromyces lactis (yeast) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.6 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Authors Authors | Murray, J. / Savva, C.G. / Shin, B.S. / Dever, T.E. / Ramakrishnan, V. / Fernandez, I.S. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Funding support |  United Kingdom, United Kingdom,  United States, 3items United States, 3items
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Elife / Year: 2016 Title: Structural characterization of ribosome recruitment and translocation by type IV IRES. Authors: Jason Murray / Christos G Savva / Byung-Sik Shin / Thomas E Dever / V Ramakrishnan / Israel S Fernández / Abstract: Viral mRNA sequences with a type IV IRES are able to initiate translation without any host initiation factors. Initial recruitment of the small ribosomal subunit as well as two translocation steps ...Viral mRNA sequences with a type IV IRES are able to initiate translation without any host initiation factors. Initial recruitment of the small ribosomal subunit as well as two translocation steps before the first peptidyl transfer are essential for the initiation of translation by these mRNAs. Using electron cryomicroscopy (cryo-EM) we have structurally characterized at high resolution how the Cricket Paralysis Virus Internal Ribosomal Entry Site (CrPV-IRES) binds the small ribosomal subunit (40S) and the translocation intermediate stabilized by elongation factor 2 (eEF2). The CrPV-IRES restricts tvhe otherwise flexible 40S head to a conformation compatible with binding the large ribosomal subunit (60S). Once the 60S is recruited, the binary CrPV-IRES/80S complex oscillates between canonical and rotated states (Fernández et al., 2014; Koh et al., 2014), as seen for pre-translocation complexes with tRNAs. Elongation factor eEF2 with a GTP analog stabilizes the ribosome-IRES complex in a rotated state with an extra ~3 degrees of rotation. Key residues in domain IV of eEF2 interact with pseudoknot I (PKI) of the CrPV-IRES stabilizing it in a conformation reminiscent of a hybrid tRNA state. The structure explains how diphthamide, a eukaryotic and archaeal specific post-translational modification of a histidine residue of eEF2, is involved in translocation. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5it7.cif.gz | 5.3 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5it7.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 5it7.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/it/5it7ftp://data.pdbj.org/pub/pdb/validation_reports/it/5it7 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  8123MC  8124C  5it9C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |




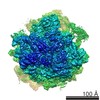
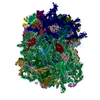
-Links
 PDBj
PDBj






























- Assembly
Assembly
| Deposited unit | 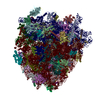
|
|---|---|
| 1 |
|
-Components
-DNA/RNA hybrid , 2 types, 2 molecules 54
| #1: DNA/RNA hybrid | Mass: 1056706.375 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) |
|---|---|
| #82: DNA/RNA hybrid | Mass: 60732.805 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Cricket paralysis virus / Production host:  |
-RNA chain , 3 types, 3 molecules 782
| #2: RNA chain | Mass: 38951.105 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: GenBank: 768843563 |
|---|---|
| #3: RNA chain | Mass: 50293.609 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: GenBank: 820806853 |
| #81: RNA chain | Mass: 579239.625 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: GenBank: 49642208 |
+Protein , 52 types, 52 molecules AACCDDEEFFGGHHIIJJMMOOPPQQRRTTUUVVYYZZaabbddeeffgghhkkmmppqq...
-60S ribosomal protein ... , 10 types, 10 molecules BBLLSSWWXXcciillnnoo
| #5: Protein | Mass: 43422.066 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CJR7 |
|---|---|
| #14: Protein | Mass: 22441.969 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q875M0 |
| #21: Protein | Mass: 20046.396 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) |
| #25: Protein | Mass: 7279.468 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: P38665 |
| #26: Protein | Mass: 13700.033 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: P48045 |
| #31: Protein | Mass: 10511.289 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: P38664 |
| #37: Protein | Mass: 10925.905 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CR18 |
| #40: Protein/peptide | Mass: 6114.286 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P04650 |
| #42: Protein/peptide | Mass: 3354.243 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P0CX86 |
| #43: Protein | Mass: 11676.897 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: P31027 |
-Ribosomal protein ... , 2 types, 2 molecules NNjj
| #16: Protein | Mass: 24193.930 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CJL7 |
|---|---|
| #38: Protein | Mass: 9679.198 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CUW0 |
-40S ribosomal protein ... , 14 types, 14 molecules ABEGIMOQVWYbcd
| #48: Protein | Mass: 22882.172 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CN12 |
|---|---|
| #49: Protein | Mass: 24488.287 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CWD0 |
| #52: Protein | Mass: 29486.312 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CWJ2 |
| #54: Protein | Mass: 25810.977 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CM04 |
| #56: Protein | Mass: 22511.531 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CMG3 |
| #60: Protein | Mass: 13124.912 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CLU4 |
| #62: Protein | Mass: 13458.439 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: P27069 |
| #64: Protein | Mass: 15656.257 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q875N2 |
| #69: Protein | Mass: 9797.949 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CXT6 |
| #70: Protein | Mass: 14513.846 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CW21 |
| #72: Protein | Mass: 15063.354 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CU44 |
| #75: Protein | Mass: 8884.362 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CNL2 |
| #76: Protein | Mass: 7073.256 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: P33285 |
| #77: Protein | Mass: 6322.149 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Kluyveromyces lactis (yeast) / References: UniProt: Q6CPG3 |
-Non-polymers , 4 types, 88 molecules 

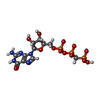

| #84: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 80 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 80 / Source method: obtained synthetically / Formula: Mg#85: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Zn#86: Chemical | ChemComp-GCP / |  Mass: 521.208 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C11H18N5O13P3 / Comment: GMP-PCP, energy-carrying molecule analogue*YM Mass: 521.208 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C11H18N5O13P3 / Comment: GMP-PCP, energy-carrying molecule analogue*YM#87: Chemical | ChemComp-6EM / ( |  Mass: 145.223 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C7H17N2O Mass: 145.223 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C7H17N2O |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 4 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||
| Buffer solution | pH: 6.5 | ||||||||||||||||||||||||||||||
| Buffer component | Conc.: 20 mM / Name: MES-KOH | ||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK II / Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 30 e/Å2 / Film or detector model: FEI FALCON II (4k x 4k) |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING ONLY | ||||||||||||
| 3D reconstruction | Resolution: 3.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 37844 / Algorithm: BACK PROJECTION / Symmetry type: POINT | ||||||||||||
| Atomic model building | Protocol: AB INITIO MODEL / Space: RECIPROCAL |
