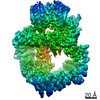
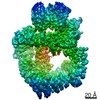
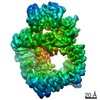
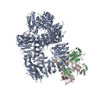
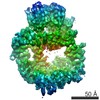
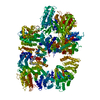
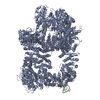
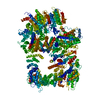
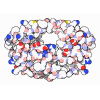
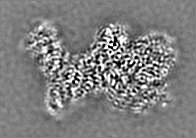
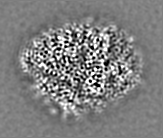
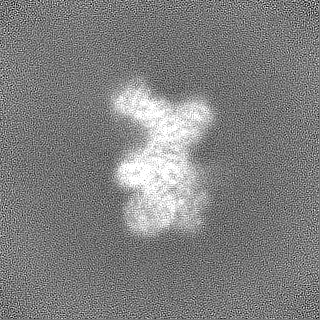
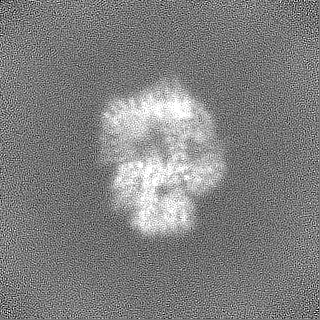
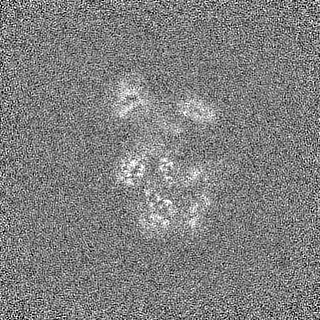
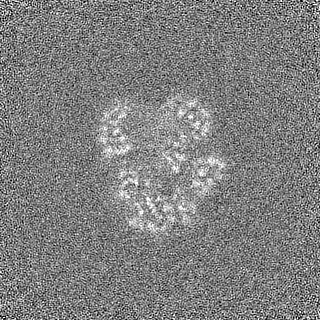
登録情報 データベース : EMDB / ID : EMD-11215タイトル Cryo-EM structure of DNA-PKcs:Ku80ct194 細胞器官・細胞要素 : DNA-PKcs in complex with Ku80 C-terminal domain細胞器官・細胞要素 : DNA-dependent protein kinase catalytic subunit,DNA-PKcsタンパク質・ペプチド : DNA-dependent protein kinase catalytic subunit,DNA-PKcs細胞器官・細胞要素 : X-ray repair cross-complementing protein 5タンパク質・ペプチド : X-ray repair cross-complementing protein 5 / / / / / / 機能・相同性 分子機能 ドメイン・相同性 構成要素
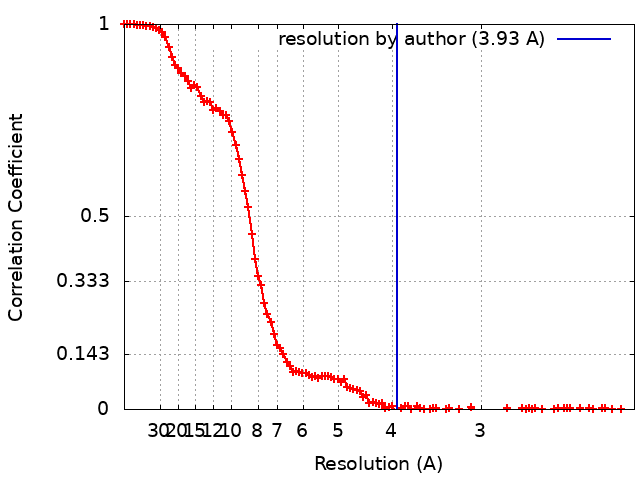
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Homo sapiens (ヒト)手法 / / 解像度 : 3.93 Å Chaplin AK / Hardwick SW 資金援助 Organization Grant number 国 Wellcome Trust 200814/Z/16/Z
ジャーナル : Nat Struct Mol Biol / 年 : 2021タイトル : Dimers of DNA-PK create a stage for DNA double-strand break repair.
著者 :
Amanda K Chaplin / Steven W Hardwick / Shikang Liang / Antonia Kefala Stavridi / Ales Hnizda / Lee R Cooper / Taiana Maia De Oliveira / Dimitri Y Chirgadze / Tom L Blundell / 要旨 :
DNA double-strand breaks are the most dangerous type of DNA damage and, if not repaired correctly, can lead to cancer. In humans, Ku70/80 recognizes DNA broken ends and recruits the DNA-dependent ... DNA double-strand breaks are the most dangerous type of DNA damage and, if not repaired correctly, can lead to cancer. In humans, Ku70/80 recognizes DNA broken ends and recruits the DNA-dependent protein kinase catalytic subunit (DNA-PKcs) to form DNA-dependent protein kinase holoenzyme (DNA-PK) in the process of non-homologous end joining (NHEJ). We present a 2.8-Å-resolution cryo-EM structure of DNA-PKcs, allowing precise amino acid sequence registration in regions uninterpreted in previous 4.3-Å X-ray maps. We also report a cryo-EM structure of DNA-PK at 3.5-Å resolution and reveal a dimer mediated by the Ku80 C terminus. Central to dimer formation is a domain swap of the conserved C-terminal helix of Ku80. Our results suggest a new mechanism for NHEJ utilizing a DNA-PK dimer to bring broken DNA ends together. Furthermore, drug inhibition of NHEJ in combination with chemo- and radiotherapy has proved successful, making these models central to structure-based drug targeting efforts. 履歴 登録 2020年6月21日 - ヘッダ(付随情報) 公開 2020年10月21日 - マップ公開 2020年10月21日 - 更新 2024年5月1日 - 現状 2024年5月1日 処理サイト : PDBe / 状態 : 公開
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード Kinase (キナーゼ) /
Kinase (キナーゼ) /  機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 英国, 1件
英国, 1件  引用
引用 構造の表示
構造の表示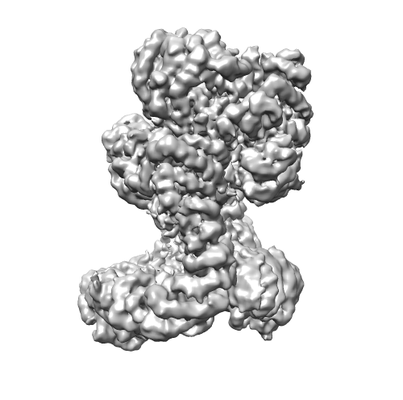
 ダウンロードとリンク
ダウンロードとリンク emd_11215.png
emd_11215.png http://ftp.pdbj.org/pub/emdb/structures/EMD-11215
http://ftp.pdbj.org/pub/emdb/structures/EMD-11215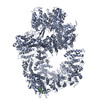




















 Z
Z Y
Y X
X






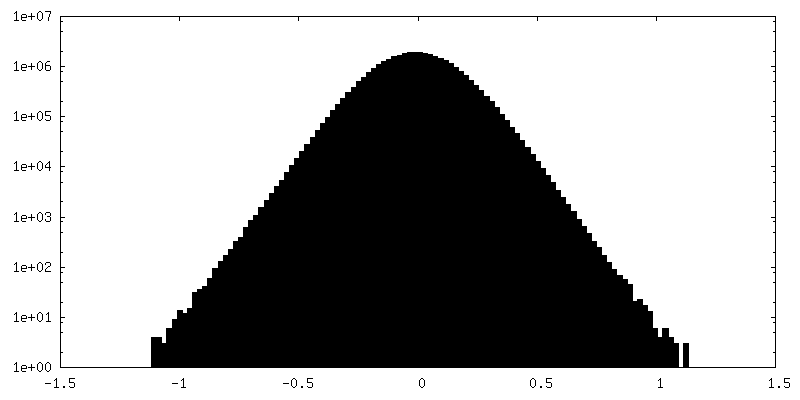
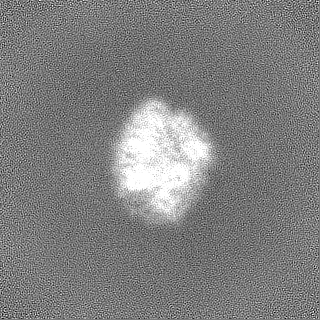
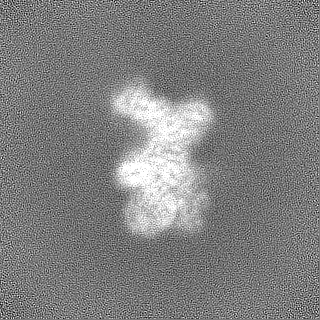
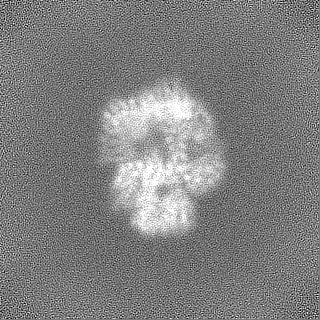




 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法