 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 3j7p | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of the 80S mammalian ribosome bound to eEF2 | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード |  RIBOSOME (リボソーム) / mammalian (哺乳類) / Sec61 (Sec61) / translocation / translation (翻訳 (生物学)) / eEF2 RIBOSOME (リボソーム) / mammalian (哺乳類) / Sec61 (Sec61) / translocation / translation (翻訳 (生物学)) / eEF2 | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報TNFR1-mediated ceramide production / Translation initiation complex formation / Formation of the ternary complex, and subsequently, the 43S complex / Ribosomal scanning and start codon recognition / SCF-beta-TrCP mediated degradation of Emi1 / Assembly of the pre-replicative complex / VLDLR internalisation and degradation / Interferon alpha/beta signaling / Negative regulators of DDX58/IFIH1 signaling / RAS processing ...TNFR1-mediated ceramide production / Translation initiation complex formation / Formation of the ternary complex, and subsequently, the 43S complex / Ribosomal scanning and start codon recognition / SCF-beta-TrCP mediated degradation of Emi1 / Assembly of the pre-replicative complex / VLDLR internalisation and degradation / Interferon alpha/beta signaling / Negative regulators of DDX58/IFIH1 signaling / RAS processing / Inactivation of CSF3 (G-CSF) signaling / Regulation of BACH1 activity / KEAP1-NFE2L2 pathway / Regulation of NF-kappa B signaling / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Translesion synthesis by REV1 / Recognition of DNA damage by PCNA-containing replication complex / Translesion Synthesis by POLH / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Downregulation of ERBB4 signaling / Spry regulation of FGF signaling / Downregulation of ERBB2:ERBB3 signaling / APC/C:Cdc20 mediated degradation of Cyclin B / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / APC-Cdc20 mediated degradation of Nek2A / EGFR downregulation / SCF(Skp2)-mediated degradation of p27/p21 / Degradation of beta-catenin by the destruction complex / TCF dependent signaling in response to WNT / Downstream TCR signaling / p75NTR recruits signalling complexes / NF-kB is activated and signals survival / Activated NOTCH1 Transmits Signal to the Nucleus / Downregulation of SMAD2/3:SMAD4 transcriptional activity / SMAD2/SMAD3:SMAD4 heterotrimer regulates transcription / Senescence-Associated Secretory Phenotype (SASP) / Stimuli-sensing channels / FCERI mediated NF-kB activation / Regulation of innate immune responses to cytosolic DNA / Autodegradation of the E3 ubiquitin ligase COP1 / Deactivation of the beta-catenin transactivating complex / ABC-family proteins mediated transport / activated TAK1 mediates p38 MAPK activation / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / AUF1 (hnRNP D0) binds and destabilizes mRNA / Asymmetric localization of PCP proteins / Degradation of AXIN / Degradation of DVL / Regulation of FZD by ubiquitination / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Regulation of TNFR1 signaling / TNFR1-induced NF-kappa-B signaling pathway / Hedgehog ligand biogenesis / CLEC7A (Dectin-1) signaling / Degradation of GLI1 by the proteasome / GLI3 is processed to GLI3R by the proteasome / Hedgehog 'on' state / Negative regulation of FGFR1 signaling / Negative regulation of FGFR2 signaling / Negative regulation of FGFR3 signaling / Negative regulation of FGFR4 signaling / Translesion synthesis by POLK / Translesion synthesis by POLI / Termination of translesion DNA synthesis / Regulation of RAS by GAPs / TNFR2 non-canonical NF-kB pathway / Negative regulation of MAPK pathway / Regulation of necroptotic cell death / MAP3K8 (TPL2)-dependent MAPK1/3 activation / HDR through Homologous Recombination (HRR) / MAPK6/MAPK4 signaling / UCH proteinases / Josephin domain DUBs / Ub-specific processing proteases / Ovarian tumor domain proteases / Metalloprotease DUBs / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / DNA Damage Recognition in GG-NER / Formation of Incision Complex in GG-NER / Gap-filling DNA repair synthesis and ligation in GG-NER / Dual Incision in GG-NER / Fanconi Anemia Pathway / Regulation of TP53 Activity through Phosphorylation / Regulation of TP53 Degradation / Regulation of TP53 Activity through Methylation / Negative regulation of MET activity / Orc1 removal from chromatin / CDK-mediated phosphorylation and removal of Cdc6 / Cyclin D associated events in G1 / G2/M Checkpoints / Ubiquitin Mediated Degradation of Phosphorylated Cdc25A / Ubiquitin-dependent degradation of Cyclin D / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / Cargo recognition for clathrin-mediated endocytosis / Downregulation of ERBB2 signaling / Synthesis of active ubiquitin: roles of E1 and E2 enzymes類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Sus scrofa (ブタ) Sus scrofa (ブタ) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.5 Å | ||||||||||||
 データ登録者 データ登録者 | Voorhees, R.M. / Fernandez, I.S. / Scheres, S.H.W. / Hegde, R.S. | ||||||||||||
 引用 引用 | ジャーナル: Cell / 年: 2014 タイトル: Structure of the mammalian ribosome-Sec61 complex to 3.4 Å resolution. 著者: Rebecca M Voorhees / Israel S Fernández / Sjors H W Scheres / Ramanujan S Hegde /  要旨: Cotranslational protein translocation is a universally conserved process for secretory and membrane protein biosynthesis. Nascent polypeptides emerging from a translating ribosome are either ...Cotranslational protein translocation is a universally conserved process for secretory and membrane protein biosynthesis. Nascent polypeptides emerging from a translating ribosome are either transported across or inserted into the membrane via the ribosome-bound Sec61 channel. Here, we report structures of a mammalian ribosome-Sec61 complex in both idle and translating states, determined to 3.4 and 3.9 Å resolution. The data sets permit building of a near-complete atomic model of the mammalian ribosome, visualization of A/P and P/E hybrid-state tRNAs, and analysis of a nascent polypeptide in the exit tunnel. Unprecedented chemical detail is observed for both the ribosome-Sec61 interaction and the conformational state of Sec61 upon ribosome binding. Comparison of the maps from idle and translating complexes suggests how conformational changes to the Sec61 channel could facilitate translocation of a secreted polypeptide. The high-resolution structure of the mammalian ribosome-Sec61 complex provides a valuable reference for future functional and structural studies. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3j7p.cif.gz | 5.4 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3j7p.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 3j7p.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j7/3j7pftp://data.pdbj.org/pub/pdb/validation_reports/j7/3j7p | HTTPS FTP |
|---|
-関連構造データ
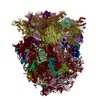

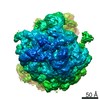

-リンク
 PDBj
PDBj































- 集合体
集合体
| 登録構造単位 | 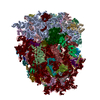
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 4種, 4分子 578S2
| #1: RNA鎖 | 28SリボソームRNA 分子量: 1187230.000 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 器官: pancreas膵臓 |
|---|---|
| #2: RNA鎖 | 5SリボソームRNA 分子量: 38691.914 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 器官: pancreas膵臓 |
| #3: RNA鎖 | 5.8SリボソームRNA 分子量: 50143.648 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 器官: pancreas膵臓 |
| #49: RNA鎖 | 分子量: 561958.812 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 器官: pancreas膵臓 |
+Ribosomal protein ... , 77種, 77分子 ABCDEFGHIJKLMNOPQRSTUVWXYZabcd...
-タンパク質 , 1種, 1分子 4
| #48: タンパク質 | EEF2 分子量: 95232.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 器官: pancreas膵臓 |
|---|
-非ポリマー , 2種, 170分子 

| #83: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 166 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 166 / 由来タイプ: 合成 / 式: Mg#84: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Zn |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: The 80S-eEF2 complex purified from porcine pancreas / タイプ: RIBOSOME |
|---|---|
| 緩衝液 | 名称: 50 mM HEPES, 200 mM potassium acetate, 15 mM magnesium acetate, 1 mM DTT, 0.25% Digitonin pH: 7.5 詳細: 50 mM HEPES, 200 mM potassium acetate, 15 mM magnesium acetate, 1 mM DTT, 0.25% Digitonin |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: Quantifoil R2/2 400 mesh copper grids |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / Temp: 120 K 詳細: 3 uL sample was incubated on the grid for 30 seconds and blotted for 9 seconds before being plunged into liquid ethane (FEI VITROBOT MARK IV). |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2014年4月7日 |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 倍率(公称値): 59000 X / 倍率(補正後): 104478 X / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 2500 nm / Cs: 2.7 mm |
| 試料ホルダ | 資料ホルダタイプ: FEI TITAN KRIOS AUTOGRID HOLDER / 温度: 70 K / 傾斜角・最大: 0 ° / 傾斜角・最小: 0 ° |
| 撮影 | 電子線照射量: 27 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) 詳細: Back-thinned |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: Each particle | |||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||
| 3次元再構成 | 手法: Single particle単粒子解析法 / 解像度: 3.5 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 36667 / ピクセルサイズ(公称値): 1.34 Å / 対称性のタイプ: POINT | |||||||||||||||
| 原子モデル構築 | B value: 37 / プロトコル: OTHER / 空間: RECIPROCAL / Target criteria: R-factor and FSC / 詳細: METHOD--Maximum likelihood | |||||||||||||||
| 原子モデル構築 |
| |||||||||||||||
| 精密化ステップ | サイクル: LAST
|

