ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9fnn | ||||||
|---|---|---|---|---|---|---|---|
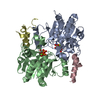
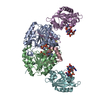
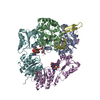
| タイトル | Cryo-EM structure of the c-di-GMP-saturated 'crown'less Bcs macrocomplex for cellulose secretion in E. coli | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | MEMBRANE PROTEIN / Bacterial cellulose secretion | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報cellulose biosynthetic process / UDP-alpha-D-glucose metabolic process / negative regulation of cell division / cytoplasmic side of plasma membrane / cell division / ATP hydrolysis activity / ATP binding / plasma membrane / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.85 Å | ||||||
 データ登録者 データ登録者 | Anso, I. / Krasteva, P.V. | ||||||
| 資金援助 | European Union, 1件
| ||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2024 タイトル: Structural basis for synthase activation and cellulose modification in the E. coli Type II Bcs secretion system. 著者: Itxaso Anso / Samira Zouhir / Thibault Géry Sana / Petya Violinova Krasteva /   要旨: Bacterial cellulosic polymers constitute a prevalent class of biofilm matrix exopolysaccharides that are synthesized by several types of bacterial cellulose secretion (Bcs) systems, which include ...Bacterial cellulosic polymers constitute a prevalent class of biofilm matrix exopolysaccharides that are synthesized by several types of bacterial cellulose secretion (Bcs) systems, which include conserved cyclic diguanylate (c-di-GMP)-dependent cellulose synthase modules together with diverse accessory subunits. In E. coli, the biogenesis of phosphoethanolamine (pEtN)-modified cellulose relies on the BcsRQABEFG macrocomplex, encompassing inner-membrane and cytosolic subunits, and an outer membrane porin, BcsC. Here, we use cryogenic electron microscopy to shed light on the molecular mechanisms of BcsA-dependent recruitment and stabilization of a trimeric BcsG pEtN-transferase for polymer modification, and a dimeric BcsF-dependent recruitment of an otherwise cytosolic BcsERQ regulatory complex. We further demonstrate that BcsE, a secondary c-di-GMP sensor, can remain dinucleotide-bound and retain the essential-for-secretion BcsRQ partners onto the synthase even in the absence of direct c-di-GMP-synthase complexation, likely lowering the threshold for c-di-GMP-dependent synthase activation. Such activation-by-proxy mechanism could allow Bcs secretion system activity even in the absence of substantial intracellular c-di-GMP increase, and is reminiscent of other widespread synthase-dependent polysaccharide secretion systems where dinucleotide sensing and/or synthase stabilization are carried out by key co-polymerase subunits. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9fnn.cif.gz | 652 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9fnn.ent.gz | 491.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9fnn.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 9fnn_validation.pdf.gz | 1.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 9fnn_full_validation.pdf.gz | 1.9 MB | 表示 | |
| XML形式データ | 9fnn_validation.xml.gz | 95.9 KB | 表示 | |
| CIF形式データ | 9fnn_validation.cif.gz | 143 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/fn/9fnnftp://data.pdbj.org/pub/pdb/validation_reports/fn/9fnn | HTTPS FTP |
-関連構造データ





-リンク
 PDBj
PDBj

- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 11分子 ABXYZEUQWRV
| #1: タンパク質 | 分子量: 104015.688 Da / 分子数: 1 / 変異: HA-FLAG-tagged at C-terminue / 由来タイプ: 組換発現 詳細: Purified Bcs macrocomplex with stoichiometry BcsA-BcsB6-BcsR2-BcsQ2-BcsE2-BcsF2-BcsG3 由来: (組換発現) | ||||||
|---|---|---|---|---|---|---|---|
| #2: タンパク質 | 分子量: 86184.383 Da / 分子数: 4 / 変異: C-terminal tail only in refined structure / 由来タイプ: 組換発現 詳細: Purified Bcs macrocomplex with stoichiometry BcsA-BcsB6-BcsR2-BcsQ2-BcsE2-BcsF2-BcsG3 由来: (組換発現) 遺伝子: bcsB, A8C65_00280, AC789_1c39010, ACU57_05345, AM464_10380, BHS81_21120, BK292_24560, BON72_13325, BON75_10020, BON76_21165, BON94_21155, BZL31_14275, C5P01_24165, C9162_26260, CI641_ ...遺伝子: bcsB, A8C65_00280, AC789_1c39010, ACU57_05345, AM464_10380, BHS81_21120, BK292_24560, BON72_13325, BON75_10020, BON76_21165, BON94_21155, BZL31_14275, C5P01_24165, C9162_26260, CI641_010915, CWS33_18410, D3O91_11715, D9I18_15765, DQF57_09480, E0J34_09745, E0K84_22715, E2127_16410, E2128_18000, E2129_18135, E2134_17800, E5P22_13645, ELV08_08610, F1E03_18495, F1E19_10130, FRV13_18850 発現宿主: #3: タンパク質 | 分子量: 60967.539 Da / 分子数: 2 / 変異: Strep-tagged at N-terminus / 由来タイプ: 組換発現 詳細: Purified Bcs macrocomplex with stoichiometry BcsA-BcsB6-BcsR2-BcsQ2-BcsE2-BcsF2-BcsG3 由来: (組換発現) #5: タンパク質 | 分子量: 27960.832 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) 遺伝子: bcsQ, yhjQ, A8C65_00290, ACU57_05335, BMT91_17060, BON75_10030, BvCmsHHP019_01723, BvCmsSINP011_05061, C2U48_15650, D3O91_11725, D9H68_14440, D9I18_15755, D9I97_13990, DAH34_22885, DAH37_ ...遺伝子: bcsQ, yhjQ, A8C65_00290, ACU57_05335, BMT91_17060, BON75_10030, BvCmsHHP019_01723, BvCmsSINP011_05061, C2U48_15650, D3O91_11725, D9H68_14440, D9I18_15755, D9I97_13990, DAH34_22885, DAH37_19450, E2127_16420, E2128_18010, E2129_18145, E2134_17810, EAI42_04085, EC1094V2_71, NCTC10429_00778, NCTC11022_03734, NCTC9058_01652 発現宿主: #6: タンパク質 | 分子量: 8645.541 Da / 分子数: 2 / 変異: His-tagged at N-terminus / 由来タイプ: 組換発現 / 由来: (組換発現) |
-Cellulose biosynthesis protein ... , 2種, 4分子 FSGD
| #4: タンパク質 | 分子量: 7378.975 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) #7: タンパク質 | 分子量: 59687.984 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: Purified Bcs macrocomplex with stoichiometry BcsA-BcsB6-BcsR2-BcsQ2-BcsE2-BcsF2-BcsG3 由来: (組換発現) |
|---|
-非ポリマー , 3種, 10分子 


| #8: 化合物 | ChemComp-C2E /  分子量: 690.411 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C20H24N10O14P2 / タイプ: SUBJECT OF INVESTIGATION 分子量: 690.411 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C20H24N10O14P2 / タイプ: SUBJECT OF INVESTIGATION#9: 化合物 |  分子量: 507.181 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM#10: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Purified Bcs macrocomplex with stoichiometry BcsA-BcsB6-BcsR2-BcsQ2-BcsE2-BcsF2-BcsG3 タイプ: COMPLEX 詳細: Locally refined c-di-GMP-bound synthase:pEtN-transferase complex from the E. coli Bcs cellulose secretion macrocomplex Entity ID: #1-#7 / 由来: RECOMBINANT |
|---|---|
| 分子量 | 値: 0.99 MDa / 実験値: NO |
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 8 詳細: 120 mM NaCL 20 mM HEPES pH8 5 mM MgCl2 10 uM ApppCp 4 uM c-di-GMP 0.01% LM-NPG |
| 試料 | 濃度: 2 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: Purification from solubilized membrane fraction using an anti-FLAG M2 affinity resin |
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: UltrAuFoil R1.2/1.3 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2100 nm / 最小 デフォーカス(公称値): 300 nm / Cs: 2.7 mm |
| 撮影 | 電子線照射量: 49.35 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 2 / 実像数: 20022 |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum LS / エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1359795 | |||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.85 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 260501 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL | |||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|