 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9eej: Crystal structure of E. coli aspartate transcarbamoylase in the R... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9eej | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of E. coli aspartate transcarbamoylase in the R-state complexed with CP, succinate, ATP, and Mg2+ | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | CYTOSOLIC PROTEIN / Complex / Allostery / ATCase / R-state / CP / succinate / ATP | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationaspartate carbamoyltransferase complex / pyrimidine nucleotide biosynthetic process / aspartate carbamoyltransferase / aspartate carbamoyltransferase activity / amino acid metabolic process / amino acid binding / 'de novo' UMP biosynthetic process / 'de novo' pyrimidine nucleobase biosynthetic process / transferase activity / metal ion binding / cytosol Similarity search - Function | ||||||||||||
| Biological species |  | ||||||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3 Å | ||||||||||||
 Authors Authors | Patterson, M.G. / Miller, R.C. / Ando, N. | ||||||||||||
| Funding support |  United States, 3items United States, 3items
| ||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2026 Title: Cooperativity in E. coli aspartate transcarbamoylase is tuned by allosteric breathing. Authors: Robert C Miller / Michael G Patterson / Neti Bhatt / Xiaokun Pei / Nozomi Ando / Abstract: Aspartate transcarbamoylase (ATCase) from Escherichia coli catalyzes a key step in pyrimidine nucleotide biosynthesis and has long served as a model for allosteric regulation. Despite decades of ...Aspartate transcarbamoylase (ATCase) from Escherichia coli catalyzes a key step in pyrimidine nucleotide biosynthesis and has long served as a model for allosteric regulation. Despite decades of study, how nucleotide binding at distant regulatory sites controls cooperativity between active sites remained unresolved. Here we show that ATCase does not simply interconvert between two conformations, as traditionally depicted, but instead samples a continuum of conformations that tune enzyme cooperativity. Using complementary cryo-electron microscopy, small-angle X-ray scattering, and crystallography under conditions that ensure full assembly of the allosteric sites, we show that ATCase behaves like a flexible balloon whose global "breathing" motions directly regulate activity: compression enforces high cooperativity, inhibiting the enzyme, whereas expansion relieves this cooperativity and activates the enzyme. We further show that all four ribonucleoside triphosphates act in symmetric pairs to tune this motion, with the pyrimidines CTP and UTP compressing the enzyme to limit further pyrimidine production, and the purines ATP and GTP expanding it to balance pyrimidine and purine pools. Together, these findings uncover a dynamic breathing mechanism for long-range allosteric communication in ATCase. #1: Journal: Acta Crystallogr D Struct Biol / Year: 2019 Title: Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Authors: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty ...Authors: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / Robert D Oeffner / Billy K Poon / Michael G Prisant / Randy J Read / Jane S Richardson / David C Richardson / Massimo D Sammito / Oleg V Sobolev / Duncan H Stockwell / Thomas C Terwilliger / Alexandre G Urzhumtsev / Lizbeth L Videau / Christopher J Williams / Paul D Adams /   Abstract: Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological ...Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological processes and to develop new therapeutics against diseases. The overall structure-solution workflow is similar for these techniques, but nuances exist because the properties of the reduced experimental data are different. Software tools for structure determination should therefore be tailored for each method. Phenix is a comprehensive software package for macromolecular structure determination that handles data from any of these techniques. Tasks performed with Phenix include data-quality assessment, map improvement, model building, the validation/rebuilding/refinement cycle and deposition. Each tool caters to the type of experimental data. The design of Phenix emphasizes the automation of procedures, where possible, to minimize repetitive and time-consuming manual tasks, while default parameters are chosen to encourage best practice. A graphical user interface provides access to many command-line features of Phenix and streamlines the transition between programs, project tracking and re-running of previous tasks. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9eej.cif.gz | 1.3 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9eej.ent.gz | 907.1 KB | Display | PDB format |
| PDBx/mmJSON format | 9eej.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ee/9eejftp://data.pdbj.org/pub/pdb/validation_reports/ee/9eej | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  9eehC  9eekC  9eelC  9eemC  9eenC  9eeoC  9eepC  9eeqC  9eerC  9eesC  9eeuC C: citing same article ( |
|---|---|
| Similar structure data |










-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| Unit cell |
|
-Components
-Protein , 2 types, 12 molecules ABCFGKDEHIJL
| #1: Protein | Mass: 34337.105 Da / Num. of mol.: 6 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: pyrB, A2J79_003077, ABE91_001635, ABT96_002053, ACN81_01160, ACW72_001368, ATM34_004868, AW118_07535, AWP47_22680, B6R15_003056, B6R31_000750, BANRA_02937, BE932_21750, BER14_11960, BG944_ ...Gene: pyrB, A2J79_003077, ABE91_001635, ABT96_002053, ACN81_01160, ACW72_001368, ATM34_004868, AW118_07535, AWP47_22680, B6R15_003056, B6R31_000750, BANRA_02937, BE932_21750, BER14_11960, BG944_000740, BGM66_001590, BGZ_01518, BGZ_05217, BJI68_10440, BK300_13470, BK383_13480, BKL28_003429, BRV02_003861, BTB68_002687, BTQ06_18960, BvCmsNSP007_04794, BVE17_001554, BXT93_08870, C0P57_000163, C1Q91_004309, C2M16_07715, C2R31_000937, C3F40_13755, C4M78_05240, C719_002693, C9114_01215, C9160_23140, C9Z68_08980, CF22_004479, CG704_05430, CIG67_22815, CLG78_001649, CQ842_08915, CQ842_12490, CR538_22700, CTR35_001468, CV628_004970, CV83915_01779, CWS33_03955, D3C88_12940, D3G36_17310, D4M65_07255, D4N09_04845, D9D43_13600, D9E49_11435, D9J61_12555, DAH27_26520, DIV22_16805, DNQ45_09630, DNX30_17140, DS732_03300, DTL43_02750, DU321_18175, E2865_05519, E5H86_07990, E6D34_03380, EAI46_26260, ECs5222, EF082_09955, EIA08_16955, EIZ93_20760, EN85_001335, EPS76_30055, EPS97_17825, EWK56_08300, ExPECSC038_04763, F7F11_04770, F7G01_01060, F7N46_16830, F9413_07035, F9B07_19120, FEH53_07480, FGAF1022_42670, FIJ20_18930, FJQ40_01260, FKO60_19415, FOI11_014380, FOI11_21300, FPI65_26085, FPS11_22335, FVB16_24685, FZU14_19575, G3V95_10555, G3W53_08345, G4A38_03455, G4A47_10535, G5603_09990, G6Z99_20225, GAI89_04005, GAJ12_10370, GKF66_08525, GNW61_17465, GNZ05_12485, GOP25_13765, GP965_09375, GP979_09965, GQE86_05410, GQM04_23140, GQM13_11160, GQM21_13835, GQN24_22415, GRC73_06815, GRW05_26905, GRW24_19535, GTP92_08545, GUC01_09445, HEP34_001710, HHH44_002259, HI055_001287, HI084_001705, HIA71_17170, HID72_003759, HIE29_003970, HJ488_000368, HJS37_003257, HKA49_000761, HLX92_11970, HLZ39_13930, HLZ50_15085, HND12_14900, HV209_29455, HVV39_15605, HVW04_12090, HVW43_13615, HVW90_11235, HVY77_22735, I6H00_15800, I6H02_17085, IFN33_000225, IG021_000549, IH772_04275, J0541_002249, J5U05_001302, J8F57_000038, KQO22_002766, NCTC10082_01748, NCTC10089_04451, NCTC10764_04395, NCTC10767_00303, NCTC10865_05523, NCTC10974_05016, NCTC11112_02870, NCTC11181_01680, NCTC11341_03161, NCTC13148_04788, NCTC7922_05337, NCTC7928_04108, NCTC8179_04755, NCTC8500_04950, NCTC8621_04481, NCTC8959_04205, NCTC8960_01943, NCTC8985_03619, NCTC9001_04952, NCTC9044_02747, NCTC9045_05133, NCTC9073_03832, NCTC9081_01672, NCTC9117_05502, NCTC9702_05190, NCTC9703_03794, OFN31_17640, OGM49_02125, P6223_000603, QDW62_23130, R8G00_26825, R8O40_003191, RZR61_03900, SAMEA3472044_01445, SAMEA3472112_04514, SAMEA3752557_01748, TUM18780_38870 Production host: #2: Protein | Mass: 17143.625 Da / Num. of mol.: 6 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: pyrI, A2J79_003078, A5U30_000934, ABE91_001630, ABT96_002052, ACN81_01165, ACU57_15645, ACW72_001369, ATM34_004869, AWP47_22675, B6R15_003055, B6R31_000751, BANRA_00251, BANRA_02936, BCB93_ ...Gene: pyrI, A2J79_003078, A5U30_000934, ABE91_001630, ABT96_002052, ACN81_01165, ACU57_15645, ACW72_001369, ATM34_004869, AWP47_22675, B6R15_003055, B6R31_000751, BANRA_00251, BANRA_02936, BCB93_002761, BE932_21745, BG944_000741, BGM66_001591, BGZ_01519, BGZ_05216, BJI68_10435, BK300_13465, BK383_13475, BKL28_003430, BMT50_05825, BRV02_003862, BTB68_002686, BTQ06_18965, BvCmsKKP061_03433, BvCmsNSP007_04795, BVE17_001555, BXT93_08875, C0P57_000164, C1Q91_004308, C2M16_07710, C2R31_000938, C3F40_13760, C4M78_05245, C7B02_12530, C9160_23135, CF22_004478, CG704_05435, CIG67_22820, CLG78_001650, CQ842_08920, CQ842_12485, CR538_22705, CTR35_001469, CV628_004971, CV83915_01780, CWS33_03950, D3C88_12935, D3G36_17315, D4M65_07260, D4N09_04840, D9D43_13595, D9E49_11440, D9J61_12550, DAH27_26525, DD762_15800, DIV22_16800, DL738_14650, DL968_06885, DNQ45_09635, DNX30_17145, DS732_03295, DTL43_02745, DU321_18180, E0I52_01830, E2865_05518, E4K51_15130, E5H86_07995, E6D34_03385, EAI46_26265, ECs5221, EF082_09960, EIA08_16950, EN85_001336, EPS97_17820, EWK56_08305, ExPECSC038_04762, F7G01_01055, F7N46_16835, F9413_07040, F9461_17650, F9B07_19115, FGAF1022_42660, FGAF848_40920, FIJ20_18925, FJQ40_01265, FKO60_19420, FOI11_014385, FOI11_21295, FPI65_26080, FPS11_22330, FTV93_15585, FWK02_19125, FZU14_19580, G3V95_10560, G3W53_08350, G4A38_03450, G4A47_10540, G5603_09985, GAI89_04010, GAJ12_10375, GNW61_17470, GOP25_13770, GP711_18320, GP954_15650, GP965_09380, GP979_09970, GQA06_14590, GQE86_05415, GQM04_23145, GQM13_11155, GQN24_22420, GQN34_01855, GRC73_06810, GRW05_26910, GRW24_19530, GUC01_09440, H0O72_05310, HEP30_009870, HEP34_001711, HHH44_002260, HI084_001706, HIA71_17175, HID72_003760, HIE29_003971, HJ488_000369, HJQ60_002439, HJS37_003258, HKA49_000762, HLZ39_13925, HLZ50_15090, HMV95_06310, HV109_21095, HV209_29450, HVV39_15600, HVW43_13620, HVW90_11240, HVY77_22740, I6H00_15805, I6H02_17080, IFC14_000343, IG021_000550, J0541_002250, J8F57_000039, JNP96_03410, KTF03_003069, NCTC10082_01749, NCTC10089_04452, NCTC10418_06552, NCTC10429_04598, NCTC10764_04396, NCTC10767_00304, NCTC10865_05524, NCTC10974_05017, NCTC11112_02869, NCTC11126_02378, NCTC11181_01681, NCTC11341_03160, NCTC13148_04787, NCTC7922_05338, NCTC7927_04891, NCTC8009_07825, NCTC8179_04754, NCTC8621_04480, NCTC8622_02187, NCTC8959_04206, NCTC8960_01944, NCTC8985_03620, NCTC9045_05134, NCTC9073_03833, NCTC9077_05555, NCTC9081_01671, NCTC9117_05503, NCTC9702_05191, NCTC9703_03795, NCTC9706_01714, NCTC9962_01701, OFN31_17645, OGM49_02120, P6223_000604, PWL68_003651, QDW62_23135, R8G00_26830, R8O40_003192, RZR61_03905, SAMEA3472044_01446, SAMEA3472112_04513, SAMEA3752557_01747, TUM18780_38880 Production host: |
|---|
-Non-polymers , 5 types, 36 molecules 
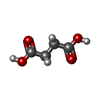



| #3: Chemical | ChemComp-CP /  Mass: 141.020 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: CH4NO5P Mass: 141.020 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: CH4NO5P#4: Chemical | ChemComp-SIN /  Mass: 118.088 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: C4H6O4 Mass: 118.088 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: C4H6O4#5: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Zn#6: Chemical | ChemComp-ATP /  Mass: 507.181 Da / Num. of mol.: 12 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 12 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM#7: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Mg |
|---|
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | N |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.23 Å3/Da / Density % sol: 61.92 % |
|---|---|
| Crystal grow | Temperature: 295 K / Method: vapor diffusion, hanging drop / pH: 6.5 Details: Protein solution: 0.016 mM ATCase in 40 mM Tris pH 7.0, 20 mM CP, 20 mM succinate, 30 mM MgCl2, and 10 mM ATP Well solution: 0.1 M Bis-Tris pH 6.5 and 20-25% PEG 300 Drops: 0.001 mL each of ...Details: Protein solution: 0.016 mM ATCase in 40 mM Tris pH 7.0, 20 mM CP, 20 mM succinate, 30 mM MgCl2, and 10 mM ATP Well solution: 0.1 M Bis-Tris pH 6.5 and 20-25% PEG 300 Drops: 0.001 mL each of protein and well solution Temp details: ambient temperature |
-Data collection
| Diffraction | Mean temperature: 100 K / Ambient temp details: cryogenic temperature / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: APS / Beamline: 24-ID-C / Wavelength: 0.9795 Å |
| Detector | Type: DECTRIS PILATUS 6M / Detector: PIXEL / Date: Mar 27, 2021 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9795 Å / Relative weight: 1 |
| Reflection | Resolution: 3→103.9 Å / Num. obs: 80502 / % possible obs: 99.3 % / Redundancy: 5.8 % / Biso Wilson estimate: 62.34 Å2 / CC1/2: 0.966 / Rmerge(I) obs: 0.2676 / Rpim(I) all: 0.1213 / Net I/σ(I): 4.2 |
| Reflection shell | Resolution: 3→3.11 Å / Rmerge(I) obs: 1.027 / Mean I/σ(I) obs: 0.7 / Num. unique obs: 7971 / CC1/2: 0.642 / Rpim(I) all: 0.454 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 3→103.9 Å / SU ML: 0.4512 / Cross valid method: FREE R-VALUE / σ(F): 1.33 / Phase error: 24.9719 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.1 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 71.23 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3→103.9 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
