 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9c68: The CRISPR associated CARF-adenosine deaminase Cad1-CARF in the c... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9c68 | ||||||
|---|---|---|---|---|---|---|---|
| Title | The CRISPR associated CARF-adenosine deaminase Cad1-CARF in the cA6 bound form | ||||||
 Components Components |
| ||||||
 Keywords Keywords | Antiviral Protein/RNA / Type-III CRISPR defense system / CARF-effector protein / adaptive immunity / deamination defense strategy / cyclic Hexa-adenylate / Antiviral Protein-RNA complex | ||||||
| Function / homology |  Function and homology information Function and homology informationadenosine deaminase / inosine biosynthetic process / hypoxanthine salvage / adenosine deaminase activity / adenosine catabolic process / cytosol Similarity search - Function | ||||||
| Biological species |  Bacteroidales bacterium (bacteria) Bacteroidales bacterium (bacteria) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.82 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.82 Å | ||||||
 Authors Authors | Majumder, P. / Patel, D.J. | ||||||
| Funding support |  United States, 1items United States, 1items
| ||||||
 Citation Citation | Journal: Cell / Year: 2024 Title: The CRISPR-associated adenosine deaminase Cad1 converts ATP to ITP to provide antiviral immunity. Authors: Christian F Baca / Puja Majumder / James H Hickling / Linzhi Ye / Marianna Teplova / Sean F Brady / Dinshaw J Patel / Luciano A Marraffini / Abstract: Type III CRISPR systems provide immunity against genetic invaders through the production of cyclic oligo-adenylate (cA) molecules that activate effector proteins that contain CRISPR-associated ...Type III CRISPR systems provide immunity against genetic invaders through the production of cyclic oligo-adenylate (cA) molecules that activate effector proteins that contain CRISPR-associated Rossman fold (CARF) domains. Here, we characterized the function and structure of an effector in which the CARF domain is fused to an adenosine deaminase domain, CRISPR-associated adenosine deaminase 1 (Cad1). We show that upon binding of cA or cA to its CARF domain, Cad1 converts ATP to ITP, both in vivo and in vitro. Cryoelectron microscopy (cryo-EM) structural studies on full-length Cad1 reveal an hexameric assembly composed of a trimer of dimers, with bound ATP at inter-domain sites required for activity and ATP/ITP within deaminase active sites. Upon synthesis of cA during phage infection, Cad1 activation leads to a growth arrest of the host that prevents viral propagation. Our findings reveal that CRISPR-Cas systems employ a wide range of molecular mechanisms beyond nucleic acid degradation to provide adaptive immunity in prokaryotes. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9c68.cif.gz | 83 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9c68.ent.gz | 60.9 KB | Display | PDB format |
| PDBx/mmJSON format | 9c68.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/c6/9c68ftp://data.pdbj.org/pub/pdb/validation_reports/c6/9c68 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  9c67C  9c69C  9c6aC  9c6cC  9c6fC  9c77C  9cdbC C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj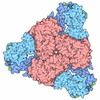
- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
| ||||||||
| Components on special symmetry positions |
|
-Components
| #1: Protein | Mass: 21069.125 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Bacteroidales bacterium (bacteria) / Gene: DCM62_02910 / Production host: #2: RNA chain | | Mass: 1930.277 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Bacteroidales bacterium (bacteria)#3: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 49 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 49 / Source method: isolated from a natural source / Formula: H2OHas protein modification | N | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.28 Å3/Da / Density % sol: 46.1 % / Description: small diamond |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 4.5 / Details: 0.1 M sodium acetate pH 4.5, 35 % (v/v) MPD |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: NSLS-II / Beamline: 17-ID-2 / Wavelength: 0.9793 Å |
| Detector | Type: DECTRIS EIGER X 9M / Detector: PIXEL / Date: Feb 4, 2024 / Details: KB bimorph mirrors |
| Radiation | Monochromator: Si(111) DCM / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9793 Å / Relative weight: 1 |
| Reflection | Resolution: 1.82→74.41 Å / Num. obs: 37752 / % possible obs: 100 % / Redundancy: 1.8 % / CC1/2: 0.999 / Rmerge(I) obs: 0.038 / Rpim(I) all: 0.038 / Rrim(I) all: 0.053 / Χ2: 0.92 / Net I/σ(I): 9.5 / Num. measured all: 69781 |
| Reflection shell | Resolution: 1.82→1.86 Å / % possible obs: 99.9 % / Redundancy: 1.9 % / Rmerge(I) obs: 2.017 / Num. measured all: 4137 / Num. unique obs: 2181 / CC1/2: 0.421 / Rpim(I) all: 2.017 / Rrim(I) all: 2.853 / Χ2: 0.79 / Net I/σ(I) obs: 0.4 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 1.82→42.96 Å / SU ML: 0.32 / Cross valid method: FREE R-VALUE / σ(F): 1.89 / Phase error: 34.89 / Stereochemistry target values: ML
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.1 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 1.82→42.96 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
