 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8twi | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
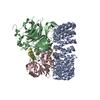
| タイトル | Cryo-EM structure of the PP2A:B55-FAM122A complex, PP2Ac body | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | SIGNALING PROTEIN / Protein Phosphatase 2A:B55 holoenzyme FAM122A inhibitor substrate binding cell cycle regulation | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報protein serine/threonine phosphatase inhibitor activity / meiotic spindle elongation / Integration of energy metabolism / PP2A-mediated dephosphorylation of key metabolic factors / RNA polymerase II CTD heptapeptide repeat S2 phosphatase activity / RNA polymerase II CTD heptapeptide repeat S7 phosphatase activity / mitotic sister chromatid separation / MASTL Facilitates Mitotic Progression / protein phosphatase type 2A complex / protein serine/threonine phosphatase complex ...protein serine/threonine phosphatase inhibitor activity / meiotic spindle elongation / Integration of energy metabolism / PP2A-mediated dephosphorylation of key metabolic factors / RNA polymerase II CTD heptapeptide repeat S2 phosphatase activity / RNA polymerase II CTD heptapeptide repeat S7 phosphatase activity / mitotic sister chromatid separation / MASTL Facilitates Mitotic Progression / protein phosphatase type 2A complex / protein serine/threonine phosphatase complex / regulation of meiotic cell cycle process involved in oocyte maturation / peptidyl-threonine dephosphorylation / meiotic sister chromatid cohesion, centromeric / INTAC complex / RNA polymerase II CTD heptapeptide repeat S5 phosphatase activity / FAR/SIN/STRIPAK complex / Regulation of glycolysis by fructose 2,6-bisphosphate metabolism / Inhibition of replication initiation of damaged DNA by RB1/E2F1 / female meiotic nuclear division / protein phosphatase regulator activity / GABA receptor binding / APC truncation mutants have impaired AXIN binding / AXIN missense mutants destabilize the destruction complex / Truncations of AMER1 destabilize the destruction complex / protein antigen binding / ERKs are inactivated / Initiation of Nuclear Envelope (NE) Reformation / positive regulation of extrinsic apoptotic signaling pathway in absence of ligand / Beta-catenin phosphorylation cascade / Signaling by GSK3beta mutants / CTNNB1 S33 mutants aren't phosphorylated / CTNNB1 S37 mutants aren't phosphorylated / CTNNB1 S45 mutants aren't phosphorylated / CTNNB1 T41 mutants aren't phosphorylated / mitotic G2/M transition checkpoint / RNA polymerase II transcription initiation surveillance / Co-stimulation by CD28 / regulation of growth / Disassembly of the destruction complex and recruitment of AXIN to the membrane / negative regulation of epithelial to mesenchymal transition / Co-inhibition by CTLA4 / protein phosphatase inhibitor activity / Platelet sensitization by LDL / protein-serine/threonine phosphatase / negative regulation of glycolytic process through fructose-6-phosphate / positive regulation of NLRP3 inflammasome complex assembly / ERK/MAPK targets / mesoderm development / protein serine/threonine phosphatase activity / vascular endothelial cell response to oscillatory fluid shear stress / T cell homeostasis / regulation of cell differentiation / regulation of G1/S transition of mitotic cell cycle / regulation of microtubule polymerization / phosphoprotein phosphatase activity / lateral plasma membrane / chromosome, centromeric region / DARPP-32 events / negative regulation of hippo signaling / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / Cyclin A/B1/B2 associated events during G2/M transition / protein dephosphorylation / spindle assembly / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Recruitment of mitotic centrosome proteins and complexes / negative regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / protein tyrosine phosphatase activity / Resolution of Sister Chromatid Cohesion / protein phosphatase 2A binding / AURKA Activation by TPX2 / Turbulent (oscillatory, disturbed) flow shear stress activates signaling by PIEZO1 and integrins in endothelial cells / Spry regulation of FGF signaling / meiotic cell cycle / chromosome segregation / Degradation of beta-catenin by the destruction complex / RHO GTPases Activate Formins / RAF activation / negative regulation of canonical Wnt signaling pathway / PKR-mediated signaling / tau protein binding / response to lead ion / Negative regulation of MAPK pathway / Cyclin D associated events in G1 / Separation of Sister Chromatids / Regulation of TP53 Degradation / spindle pole / Regulation of PLK1 Activity at G2/M Transition / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / mitotic cell cycle / microtubule cytoskeleton / PI5P, PP2A and IER3 Regulate PI3K/AKT Signaling / positive regulation of cell growth / protein-containing complex assembly / neuron projection 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.69 Å | ||||||||||||
 データ登録者 データ登録者 | Fuller, J.R. / Padi, S.K.R. / Peti, W. / Page, R. | ||||||||||||
| 資金援助 |  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2024 タイトル: Cryo-EM structures of PP2A:B55-FAM122A and PP2A:B55-ARPP19. 著者: Sathish K R Padi / Margaret R Vos / Rachel J Godek / James R Fuller / Thomas Kruse / Jamin B Hein / Jakob Nilsson / Matthew S Kelker / Rebecca Page / Wolfgang Peti /  要旨: Progression through the cell cycle is controlled by regulated and abrupt changes in phosphorylation. Mitotic entry is initiated by increased phosphorylation of mitotic proteins, a process driven by ...Progression through the cell cycle is controlled by regulated and abrupt changes in phosphorylation. Mitotic entry is initiated by increased phosphorylation of mitotic proteins, a process driven by kinases, whereas mitotic exit is achieved by counteracting dephosphorylation, a process driven by phosphatases, especially PP2A:B55. Although the role of kinases in mitotic entry is well established, recent data have shown that mitosis is only successfully initiated when the counterbalancing phosphatases are also inhibited. Inhibition of PP2A:B55 is achieved by the intrinsically disordered proteins ARPP19 and FAM122A. Despite their critical roles in mitosis, the mechanisms by which they achieve PP2A:B55 inhibition is unknown. Here, we report the single-particle cryo-electron microscopy structures of PP2A:B55 bound to phosphorylated ARPP19 and FAM122A. Consistent with our complementary NMR spectroscopy studies, both intrinsically disordered proteins bind PP2A:B55, but do so in highly distinct manners, leveraging multiple distinct binding sites on B55. Our extensive structural, biophysical and biochemical data explain how substrates and inhibitors are recruited to PP2A:B55 and provide a molecular roadmap for the development of therapeutic interventions for PP2A:B55-related diseases. #1: ジャーナル: Acta Crystallogr D Struct Biol / 年: 2018 タイトル: Real-space refinement in PHENIX for cryo-EM and crystallography. 著者: Pavel V Afonine / Billy K Poon / Randy J Read / Oleg V Sobolev / Thomas C Terwilliger / Alexandre Urzhumtsev / Paul D Adams /   要旨: This article describes the implementation of real-space refinement in the phenix.real_space_refine program from the PHENIX suite. The use of a simplified refinement target function enables very fast ...This article describes the implementation of real-space refinement in the phenix.real_space_refine program from the PHENIX suite. The use of a simplified refinement target function enables very fast calculation, which in turn makes it possible to identify optimal data-restraint weights as part of routine refinements with little runtime cost. Refinement of atomic models against low-resolution data benefits from the inclusion of as much additional information as is available. In addition to standard restraints on covalent geometry, phenix.real_space_refine makes use of extra information such as secondary-structure and rotamer-specific restraints, as well as restraints or constraints on internal molecular symmetry. The re-refinement of 385 cryo-EM-derived models available in the Protein Data Bank at resolutions of 6 Å or better shows significant improvement of the models and of the fit of these models to the target maps. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8twi.cif.gz | 215.5 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8twi.ent.gz | 166.4 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8twi.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8twi_validation.pdf.gz | 1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8twi_full_validation.pdf.gz | 1 MB | 表示 | |
| XML形式データ | 8twi_validation.xml.gz | 39.8 KB | 表示 | |
| CIF形式データ | 8twi_validation.cif.gz | 56.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/tw/8twiftp://data.pdbj.org/pub/pdb/validation_reports/tw/8twi | HTTPS FTP |
-関連構造データ





-リンク
 PDBj
PDBj
















- 集合体
集合体
| 登録構造単位 | 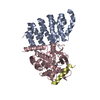
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 64957.980 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PPP2R1A / 発現宿主:  |
|---|---|
| #2: タンパク質 | 分子量: 35845.375 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PPP2CA / 発現宿主: Homo sapiens (ヒト)参照: UniProt: P67775, protein-serine/threonine phosphatase |
| #3: タンパク質 | 分子量: 10680.907 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PABIR1, C9orf42, FAM122A / 発現宿主: |
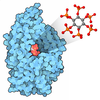
| #4: 化合物 | ChemComp-FE /   分子量: 55.845 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Fe / タイプ: SUBJECT OF INVESTIGATION 分子量: 55.845 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Fe / タイプ: SUBJECT OF INVESTIGATION |
| #5: 化合物 | ChemComp-ZN /   分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION |
| 研究の焦点であるリガンドがあるか | Y |
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Quadruple complex of PP2A:B55 (PP2Aa:PP2Ac:B55) bound to FAM122A タイプ: COMPLEX / Entity ID: #1-#3 / 由来: MULTIPLE SOURCES | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 8 詳細: CHAPSO was added only immediately prior to vitrification | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 1.2 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 291 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 1900 nm / 最小 デフォーカス(公称値): 700 nm / Calibrated defocus min: 550 nm / 最大 デフォーカス(補正後): 2530 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: ZEMLIN TABLEAU |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 70 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 詳細: Camera was operated in CDS mode, writing super-resolution movies with 59 frames |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum / エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1248538 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.69 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 103522 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL / Target criteria: Cross-correlation 詳細: Iterating between manual refinement in Coot and automated real-space refinement in Phenix | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|

