 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8so0 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of the PP2A:B55-FAM122A complex | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | SIGNALING PROTEIN / Protein Phosphatase 2A:B55 holoenzyme FAM122A inhibitor substrate binding cell cycle regulation | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationprotein serine/threonine phosphatase inhibitor activity / regulation of chromosome segregation / meiotic spindle elongation / PP2A-mediated dephosphorylation of key metabolic factors / RNA polymerase II CTD heptapeptide repeat S2 phosphatase activity / RNA polymerase II CTD heptapeptide repeat S7 phosphatase activity / peptidyl-threonine dephosphorylation / regulation of meiotic cell cycle process involved in oocyte maturation / mitotic sister chromatid separation / MASTL Facilitates Mitotic Progression ...protein serine/threonine phosphatase inhibitor activity / regulation of chromosome segregation / meiotic spindle elongation / PP2A-mediated dephosphorylation of key metabolic factors / RNA polymerase II CTD heptapeptide repeat S2 phosphatase activity / RNA polymerase II CTD heptapeptide repeat S7 phosphatase activity / peptidyl-threonine dephosphorylation / regulation of meiotic cell cycle process involved in oocyte maturation / mitotic sister chromatid separation / MASTL Facilitates Mitotic Progression / protein phosphatase type 2A complex / meiotic sister chromatid cohesion, centromeric / INTAC complex / RNA polymerase II CTD heptapeptide repeat S5 phosphatase activity / FAR/SIN/STRIPAK complex / female meiotic nuclear division / Regulation of glycolysis by fructose 2,6-bisphosphate metabolism / Inhibition of replication initiation of damaged DNA by RB1/E2F1 / protein phosphatase regulator activity / protein antigen binding / GABA receptor binding / APC truncation mutants have impaired AXIN binding / AXIN missense mutants destabilize the destruction complex / Truncations of AMER1 destabilize the destruction complex / positive regulation of extrinsic apoptotic signaling pathway in absence of ligand / ERKs are inactivated / mitotic G2/M transition checkpoint / Initiation of Nuclear Envelope (NE) Reformation / Beta-catenin phosphorylation cascade / Signaling by GSK3beta mutants / CTNNB1 S33 mutants aren't phosphorylated / CTNNB1 S37 mutants aren't phosphorylated / CTNNB1 S45 mutants aren't phosphorylated / CTNNB1 T41 mutants aren't phosphorylated / Co-stimulation by CD28 / RNA polymerase II transcription initiation surveillance / regulation of growth / Disassembly of the destruction complex and recruitment of AXIN to the membrane / protein dephosphorylation / negative regulation of epithelial to mesenchymal transition / Co-inhibition by CTLA4 / Platelet sensitization by LDL / protein phosphatase inhibitor activity / protein-serine/threonine phosphatase / response to morphine / negative regulation of glycolytic process through fructose-6-phosphate / ERK/MAPK targets / T cell homeostasis / vascular endothelial cell response to oscillatory fluid shear stress / mesoderm development / protein serine/threonine phosphatase activity / positive regulation of NLRP3 inflammasome complex assembly / regulation of cell differentiation / regulation of microtubule polymerization / lateral plasma membrane / chromosome, centromeric region / DARPP-32 events / enzyme-substrate adaptor activity / negative regulation of hippo signaling / Cyclin A/B1/B2 associated events during G2/M transition / regulation of G1/S transition of mitotic cell cycle / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / spindle assembly / phosphoprotein phosphatase activity / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Recruitment of mitotic centrosome proteins and complexes / protein tyrosine phosphatase activity / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / protein phosphatase 2A binding / Turbulent (oscillatory, disturbed) flow shear stress activates signaling by PIEZO1 and integrins in endothelial cells / AURKA Activation by TPX2 / Resolution of Sister Chromatid Cohesion / negative regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / chromosome segregation / negative regulation of canonical Wnt signaling pathway / RAF activation / RHO GTPases Activate Formins / meiotic cell cycle / Spry regulation of FGF signaling / PKR-mediated signaling / response to lead ion / tau protein binding / Degradation of beta-catenin by the destruction complex / spindle pole / Cyclin D associated events in G1 / Negative regulation of MAPK pathway / Separation of Sister Chromatids / Regulation of TP53 Degradation / Regulation of PLK1 Activity at G2/M Transition / mitotic cell cycle / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / microtubule cytoskeleton / PI5P, PP2A and IER3 Regulate PI3K/AKT Signaling / positive regulation of cell growth / protein-containing complex assembly Similarity search - Function | ||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.79961 Å | ||||||||||||
 Authors Authors | Fuller, J.R. / Padi, S.K.R. / Peti, W. / Page, R. | ||||||||||||
| Funding support |  United States, 3items United States, 3items
| ||||||||||||
 Citation Citation | Journal: Nature / Year: 2024 Title: Cryo-EM structures of PP2A:B55-FAM122A and PP2A:B55-ARPP19. Authors: Sathish K R Padi / Margaret R Vos / Rachel J Godek / James R Fuller / Thomas Kruse / Jamin B Hein / Jakob Nilsson / Matthew S Kelker / Rebecca Page / Wolfgang Peti /  Abstract: Progression through the cell cycle is controlled by regulated and abrupt changes in phosphorylation. Mitotic entry is initiated by increased phosphorylation of mitotic proteins, a process driven by ...Progression through the cell cycle is controlled by regulated and abrupt changes in phosphorylation. Mitotic entry is initiated by increased phosphorylation of mitotic proteins, a process driven by kinases, whereas mitotic exit is achieved by counteracting dephosphorylation, a process driven by phosphatases, especially PP2A:B55. Although the role of kinases in mitotic entry is well established, recent data have shown that mitosis is only successfully initiated when the counterbalancing phosphatases are also inhibited. Inhibition of PP2A:B55 is achieved by the intrinsically disordered proteins ARPP19 and FAM122A. Despite their critical roles in mitosis, the mechanisms by which they achieve PP2A:B55 inhibition is unknown. Here, we report the single-particle cryo-electron microscopy structures of PP2A:B55 bound to phosphorylated ARPP19 and FAM122A. Consistent with our complementary NMR spectroscopy studies, both intrinsically disordered proteins bind PP2A:B55, but do so in highly distinct manners, leveraging multiple distinct binding sites on B55. Our extensive structural, biophysical and biochemical data explain how substrates and inhibitors are recruited to PP2A:B55 and provide a molecular roadmap for the development of therapeutic interventions for PP2A:B55-related diseases. #1: Journal: Acta Crystallogr D Struct Biol / Year: 2018 Title: Real-space refinement in PHENIX for cryo-EM and crystallography. Authors: Pavel V Afonine / Billy K Poon / Randy J Read / Oleg V Sobolev / Thomas C Terwilliger / Alexandre Urzhumtsev / Paul D Adams /   Abstract: This article describes the implementation of real-space refinement in the phenix.real_space_refine program from the PHENIX suite. The use of a simplified refinement target function enables very fast ...This article describes the implementation of real-space refinement in the phenix.real_space_refine program from the PHENIX suite. The use of a simplified refinement target function enables very fast calculation, which in turn makes it possible to identify optimal data-restraint weights as part of routine refinements with little runtime cost. Refinement of atomic models against low-resolution data benefits from the inclusion of as much additional information as is available. In addition to standard restraints on covalent geometry, phenix.real_space_refine makes use of extra information such as secondary-structure and rotamer-specific restraints, as well as restraints or constraints on internal molecular symmetry. The re-refinement of 385 cryo-EM-derived models available in the Protein Data Bank at resolutions of 6 Å or better shows significant improvement of the models and of the fit of these models to the target maps. | ||||||||||||
| History |
|
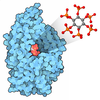
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8so0.cif.gz | 639.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8so0.ent.gz | 422 KB | Display | PDB format |
| PDBx/mmJSON format | 8so0.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/so/8so0ftp://data.pdbj.org/pub/pdb/validation_reports/so/8so0 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  40644MC  8ttbC  8tweC  8twiC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj
















- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Serine/threonine-protein phosphatase 2A ... , 3 types, 3 molecules ABC
| #1: Protein | Mass: 64957.980 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PPP2R1A / Production host:  |
|---|---|
| #2: Protein | Mass: 52101.340 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PPP2R2A / Production host: Homo sapiens (human) / References: UniProt: P63151 |
| #3: Protein | Mass: 35845.375 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PPP2CA / Production host: Homo sapiens (human)References: UniProt: P67775, protein-serine/threonine phosphatase |
-Protein , 1 types, 1 molecules D
| #4: Protein | Mass: 10680.907 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PABIR1, C9orf42, FAM122A / Production host: |
|---|
-Non-polymers , 2 types, 2 molecules 

| #5: Chemical | ChemComp-FE /  Mass: 55.845 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Fe Mass: 55.845 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Fe |
|---|---|
| #6: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn |
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Quadruple complex of PP2A:B55 (PP2Aa:PP2Ac:B55) bound to FAM122A Type: COMPLEX / Entity ID: #1-#4 / Source: MULTIPLE SOURCES | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Experimental value: NO | ||||||||||||||||||||||||||||||
| Buffer solution | pH: 8 Details: CHAPSO was added only immediately prior to vitrification | ||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||
| Specimen | Conc.: 1.2 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Specimen support | Details: Gatan Solarus, 20W power for 35s using room air / Grid material: GOLD / Grid mesh size: 300 divisions/in. / Grid type: UltrAuFoil R1.2/1.3 | ||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 291 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 81000 X / Nominal defocus max: 1900 nm / Nominal defocus min: 700 nm / Calibrated defocus min: 550 nm / Calibrated defocus max: 2530 nm / Cs: 2.7 mm / C2 aperture diameter: 50 µm / Alignment procedure: ZEMLIN TABLEAU |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 70 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Num. of grids imaged: 1 Details: Camera was operated in CDS mode, writing super-resolution movies with 59 frames |
| EM imaging optics | Energyfilter name: GIF Bioquantum / Energyfilter slit width: 20 eV |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 1248538 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.79961 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 25000 / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: Cross-correlation Details: Iterating between manual refinement in Coot and automated real-space refinement in Phenix | ||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Cross valid method: NONE Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 78.92 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|

