ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
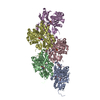
| 登録情報 | データベース: PDB / ID: 8rtt | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of the formin Cdc12 bound to the barbed end of phalloidin-stabilized F-actin. | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | STRUCTURAL PROTEIN / actin / formin / Cdc12 / actin assembly. | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報protein localization to mitotic actomyosin contractile ring / F-bar domain binding / medial cortical node / mitotic actomyosin contractile ring, proximal layer / mitotic actomyosin contractile ring / medial cortex / mitotic actomyosin contractile ring assembly / positive regulation of norepinephrine uptake / cellular response to cytochalasin B / bBAF complex ...protein localization to mitotic actomyosin contractile ring / F-bar domain binding / medial cortical node / mitotic actomyosin contractile ring, proximal layer / mitotic actomyosin contractile ring / medial cortex / mitotic actomyosin contractile ring assembly / positive regulation of norepinephrine uptake / cellular response to cytochalasin B / bBAF complex / npBAF complex / nBAF complex / brahma complex / regulation of transepithelial transport / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / GBAF complex / Formation of annular gap junctions / Formation of the dystrophin-glycoprotein complex (DGC) / protein localization to adherens junction / Gap junction degradation / regulation of G0 to G1 transition / Folding of actin by CCT/TriC / Cell-extracellular matrix interactions / dense body / barbed-end actin filament capping / postsynaptic actin cytoskeleton / Tat protein binding / mating projection tip / Prefoldin mediated transfer of substrate to CCT/TriC / RSC-type complex / regulation of nucleotide-excision repair / regulation of double-strand break repair / adherens junction assembly / Adherens junctions interactions / RHOF GTPase cycle / apical protein localization / Sensory processing of sound by outer hair cells of the cochlea / tight junction / Interaction between L1 and Ankyrins / SWI/SNF complex / regulation of mitotic metaphase/anaphase transition / Sensory processing of sound by inner hair cells of the cochlea / positive regulation of T cell differentiation / apical junction complex / regulation of norepinephrine uptake / positive regulation of double-strand break repair / transporter regulator activity / cell division site / maintenance of blood-brain barrier / nitric-oxide synthase binding / cortical cytoskeleton / establishment or maintenance of cell polarity / NuA4 histone acetyltransferase complex / positive regulation of stem cell population maintenance / Regulation of MITF-M-dependent genes involved in pigmentation / Recycling pathway of L1 / brush border / regulation of G1/S transition of mitotic cell cycle / actin filament bundle assembly / kinesin binding / negative regulation of cell differentiation / EPH-ephrin mediated repulsion of cells / regulation of synaptic vesicle endocytosis / RHO GTPases Activate WASPs and WAVEs / positive regulation of myoblast differentiation / RHO GTPases activate IQGAPs / regulation of protein localization to plasma membrane / positive regulation of double-strand break repair via homologous recombination / actin filament polymerization / cytoskeleton organization / EPHB-mediated forward signaling / substantia nigra development / axonogenesis / calyx of Held / nitric-oxide synthase regulator activity / DNA Damage Recognition in GG-NER / adherens junction / actin filament / Translocation of SLC2A4 (GLUT4) to the plasma membrane / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / FCGR3A-mediated phagocytosis / positive regulation of cell differentiation / cell motility / RHO GTPases Activate Formins / Signaling by high-kinase activity BRAF mutants / MAP2K and MAPK activation / kinetochore / Regulation of actin dynamics for phagocytic cup formation / B-WICH complex positively regulates rRNA expression / structural constituent of cytoskeleton / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / small GTPase binding / platelet aggregation / VEGFA-VEGFR2 Pathway / tau protein binding / Schaffer collateral - CA1 synapse / nuclear matrix / cytoplasmic ribonucleoprotein granule / UCH proteinases 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)  Amanita phalloides (タマゴテングタケ) Amanita phalloides (タマゴテングタケ) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.56 Å | ||||||||||||
 データ登録者 データ登録者 | Oosterheert, W. / Boiero Sanders, M. / Funk, J. / Prumbaum, D. / Raunser, S. / Bieling, P. | ||||||||||||
| 資金援助 |  ドイツ, European Union, 3件 ドイツ, European Union, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Science / 年: 2024 タイトル: Molecular mechanism of actin filament elongation by formins. 著者: Wout Oosterheert / Micaela Boiero Sanders / Johanna Funk / Daniel Prumbaum / Stefan Raunser / Peter Bieling / 要旨: Formins control the assembly of actin filaments (F-actin) that drive cell morphogenesis and motility in eukaryotes. However, their molecular interaction with F-actin and their mechanism of action ...Formins control the assembly of actin filaments (F-actin) that drive cell morphogenesis and motility in eukaryotes. However, their molecular interaction with F-actin and their mechanism of action remain unclear. In this work, we present high-resolution cryo-electron microscopy structures of F-actin barbed ends bound by three distinct formins, revealing a common asymmetric formin conformation imposed by the filament. Formation of new intersubunit contacts during actin polymerization sterically displaces formin and triggers its translocation. This "undock-and-lock" mechanism explains how actin-filament growth is coordinated with formin movement. Filament elongation speeds are controlled by the positioning and stability of actin-formin interfaces, which distinguish fast and slow formins. Furthermore, we provide a structure of the actin-formin-profilin ring complex, which resolves how profilin is rapidly released from the barbed end during filament elongation. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8rtt.cif.gz | 404.3 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8rtt.ent.gz | 329.9 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8rtt.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8rtt_validation.pdf.gz | 1.5 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8rtt_full_validation.pdf.gz | 1.5 MB | 表示 | |
| XML形式データ | 8rtt_validation.xml.gz | 74.5 KB | 表示 | |
| CIF形式データ | 8rtt_validation.cif.gz | 108.6 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/rt/8rttftp://data.pdbj.org/pub/pdb/validation_reports/rt/8rtt | HTTPS FTP |
-関連構造データ







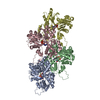

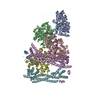
-リンク
 PDBj
PDBj
















- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 2種, 6分子 ABCDEF
| #1: タンパク質 | 分子量: 41763.617 Da / 分子数: 4 / 変異: C272A / 由来タイプ: 組換発現 詳細: Recombinant beta-actin purified from BTI-Tnao38 cells. 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ACTB / プラスミド: p2336 pFL_ACTB_C272A / 細胞株 (発現宿主): BTI-Tnao38 / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P60709 Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P60709#2: タンパク質 | 分子量: 48495.242 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: FH2 domain of S. pombe Cdc12, purified from E. coli cells. 由来: (組換発現) 遺伝子: cdc12, SPAC1F5.04c / プラスミド: pETM11-SUMO3 / 詳細 (発現宿主): Contains N-terminal His10-Sumo tag. / 発現宿主:  |
|---|
-タンパク質・ペプチド , 1種, 3分子 HIJ
| #3: タンパク質・ペプチド | タイプ: Peptide-like / クラス: 毒素 / 分子量: 808.899 Da / 分子数: 3 / 由来タイプ: 天然 / 詳細: phalloidin, purified from Amanita phalloides. 由来: (天然) Amanita phalloides (タマゴテングタケ)Plasmid details: Bought from sigma. / 参照: BIRD: PRD_002366 |
|---|
-非ポリマー , 3種, 11分子 

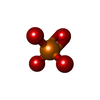
| #4: 化合物 | ChemComp-ADP /  分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM#5: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg#6: 化合物 |  分子量: 94.971 Da / 分子数: 3 / 由来タイプ: 合成 / 式: PO4 分子量: 94.971 Da / 分子数: 3 / 由来タイプ: 合成 / 式: PO4 |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | |||||||||||||||||||||||||||||||||||
| 由来(天然) |
| |||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| |||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.1 詳細: 1xKMEH (10 mM HEPES pH 7.1, 100 mM KCl, 2 mM MgCl2, 1 mM EGTA, 0.5 mM TCEP) | |||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R2/1 | |||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE-PROPANE / 湿度: 100 % / 凍結前の試料温度: 286 K / 詳細: 3 seconds, force 0. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS 詳細: 300 kV Titan Krios G2 microscope (Thermo Fisher Scientific) with an in-column Cs-corrector. |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 2700 nm / 最小 デフォーカス(公称値): 1200 nm / Cs: 0.01 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 64.6 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 20393 |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum / 詳細: Gatan energy filter. / エネルギーフィルタースリット幅: 15 eV 球面収差補正装置: Titan Krios G2 microscope (Thermo Fisher Scientific) with an in-column Cs-corrector. |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 5200600 / 詳細: Particles picked using SPHIRE-crYOLO. | ||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.56 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 154570 / 詳細: Refinement performed in RELION. / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 3D fitting-ID: 1
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 63.41 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|