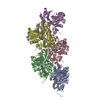
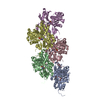
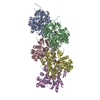
Journal: Nature / Year: 2022 Title: Structural basis of actin filament assembly and aging. Authors: Wout Oosterheert / Björn U Klink / Alexander Belyy / Sabrina Pospich / Stefan Raunser / Abstract: The dynamic turnover of actin filaments (F-actin) controls cellular motility in eukaryotes and is coupled to changes in the F-actin nucleotide state. It remains unclear how F-actin hydrolyses ATP and ...The dynamic turnover of actin filaments (F-actin) controls cellular motility in eukaryotes and is coupled to changes in the F-actin nucleotide state. It remains unclear how F-actin hydrolyses ATP and subsequently undergoes subtle conformational rearrangements that ultimately lead to filament depolymerization by actin-binding proteins. Here we present cryo-electron microscopy structures of F-actin in all nucleotide states, polymerized in the presence of Mg or Ca at approximately 2.2 Å resolution. The structures show that actin polymerization induces the relocation of water molecules in the nucleotide-binding pocket, activating one of them for the nucleophilic attack of ATP. Unexpectedly, the back door for the subsequent release of inorganic phosphate (P) is closed in all structures, indicating that P release occurs transiently. The small changes in the nucleotide-binding pocket after ATP hydrolysis and P release are sensed by a key amino acid, amplified and transmitted to the filament periphery. Furthermore, differences in the positions of water molecules in the nucleotide-binding pocket explain why Ca-actin shows slower polymerization rates than Mg-actin. Our work elucidates the solvent-driven rearrangements that govern actin filament assembly and aging and lays the foundation for the rational design of drugs and small molecules for imaging and therapeutic applications.
History
Deposition
Jun 6, 2022
Deposition site: PDBE / Processing site: PDBE
Revision 1.0
Aug 10, 2022
Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: EM metadata / Data content type: EM metadata / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Additional map / Data content type: Additional map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: FSC / Data content type: FSC / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Half map / Part number: 1 / Data content type: Half map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Half map / Part number: 2 / Data content type: Half map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Image / Data content type: Image / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Mask / Data content type: Mask / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Primary map / Data content type: Primary map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Additional map / Data content type: Additional map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: FSC / Data content type: FSC / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Half map / Part number: 1 / Data content type: Half map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Half map / Part number: 2 / Data content type: Half map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Image / Data content type: Image / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Mask / Data content type: Mask / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Primary map / Data content type: Primary map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Additional map / Data content type: Additional map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: FSC / Data content type: FSC / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Half map / Part number: 1 / Data content type: Half map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Half map / Part number: 2 / Data content type: Half map / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Image / Data content type: Image / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Mask / Data content type: Mask / Provider: repository / Type: Initial release
Revision 1.0
Aug 10, 2022
Data content type: Primary map / Data content type: Primary map / Provider: repository / Type: Initial release
Data content type: EM metadata / Data content type: EM metadata / EM metadata / EM metadata Group: Data processing / Experimental summary / Refinement description Data content type: EM metadata / EM metadata / EM metadata Category: em_3d_fitting_list / em_admin / pdbx_initial_refinement_model Data content type: EM metadata / EM metadata ...EM metadata / EM metadata / EM metadata / EM metadata / EM metadata Item: _em_3d_fitting_list.accession_code / _em_3d_fitting_list.initial_refinement_model_id ..._em_3d_fitting_list.accession_code / _em_3d_fitting_list.initial_refinement_model_id / _em_3d_fitting_list.source_name / _em_3d_fitting_list.type / _em_admin.last_update
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
 Authors
Authors Germany, 4items
Germany, 4items  Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads







 PDBj
PDBj


 Assembly
Assembly