 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8rg0: Structure of human eIF3 core from closed 48S translation initiati... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8rg0 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of human eIF3 core from closed 48S translation initiation complex | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | RIBOSOME / TRANSLATION / initiation / 48S / eIF / human / eukaryotic / factor / codon / scanning / open / reading | |||||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of mRNA binding / viral translational termination-reinitiation / eukaryotic translation initiation factor 3 complex, eIF3e / cap-dependent translational initiation / eukaryotic translation initiation factor 3 complex, eIF3m / IRES-dependent viral translational initiation / translation reinitiation / eukaryotic translation initiation factor 3 complex / formation of cytoplasmic translation initiation complex / cytoplasmic translational initiation ...positive regulation of mRNA binding / viral translational termination-reinitiation / eukaryotic translation initiation factor 3 complex, eIF3e / cap-dependent translational initiation / eukaryotic translation initiation factor 3 complex, eIF3m / IRES-dependent viral translational initiation / translation reinitiation / eukaryotic translation initiation factor 3 complex / formation of cytoplasmic translation initiation complex / cytoplasmic translational initiation / multi-eIF complex / eukaryotic 43S preinitiation complex / mRNA cap binding / eukaryotic 48S preinitiation complex / translation at postsynapse / negative regulation of RNA splicing / metal-dependent deubiquitinase activity / regulation of translational initiation / nuclear-transcribed mRNA catabolic process, nonsense-mediated decay / translation at presynapse / Formation of the ternary complex, and subsequently, the 43S complex / erythrocyte homeostasis / rRNA modification in the nucleus and cytosol / cytoplasmic side of rough endoplasmic reticulum membrane / laminin receptor activity / Ribosomal scanning and start codon recognition / Translation initiation complex formation / SARS-CoV-1 modulates host translation machinery / Peptide chain elongation / Selenocysteine synthesis / Formation of a pool of free 40S subunits / Eukaryotic Translation Termination / Response of EIF2AK4 (GCN2) to amino acid deficiency / SRP-dependent cotranslational protein targeting to membrane / Viral mRNA Translation / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / GTP hydrolysis and joining of the 60S ribosomal subunit / L13a-mediated translational silencing of Ceruloplasmin expression / Major pathway of rRNA processing in the nucleolus and cytosol / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / laminin binding / translation regulator activity / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / translation initiation factor binding / negative regulation of translational initiation / negative regulation of proteasomal ubiquitin-dependent protein catabolic process / cytosolic ribosome / Mitotic Prometaphase / antiviral innate immune response / EML4 and NUDC in mitotic spindle formation / translation initiation factor activity / Resolution of Sister Chromatid Cohesion / erythrocyte differentiation / positive regulation of translation / maturation of SSU-rRNA / translational initiation / small-subunit processome / RHO GTPases Activate Formins / PML body / mRNA 5'-UTR binding / Regulation of expression of SLITs and ROBOs / fibrillar center / metallopeptidase activity / rRNA processing / Separation of Sister Chromatids / ribosome biogenesis / presynapse / ribosome binding / virus receptor activity / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / cytosolic small ribosomal subunit / small ribosomal subunit rRNA binding / SARS-CoV-2 modulates host translation machinery / ubiquitinyl hydrolase 1 / cysteine-type deubiquitinase activity / cytoplasmic translation / cell differentiation / postsynaptic density / ribosome / structural constituent of ribosome / cadherin binding / translation / ribonucleoprotein complex / focal adhesion / mRNA binding / synapse / negative regulation of apoptotic process / chromatin / nucleolus / structural molecule activity / negative regulation of transcription by RNA polymerase II / endoplasmic reticulum / proteolysis / DNA binding / RNA binding / extracellular exosome / zinc ion binding / nucleoplasm Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.4 Å | |||||||||
 Authors Authors | Petrychenko, V. / Yi, S.-H. / Liedtke, D. / Peng, B.Z. / Rodnina, M.V. / Fischer, N. | |||||||||
| Funding support |  Germany, 2items Germany, 2items
| |||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2025 Title: Structural basis for translational control by the human 48S initiation complex. Authors: Valentyn Petrychenko / Sung-Hui Yi / David Liedtke / Bee-Zen Peng / Marina V Rodnina / Niels Fischer / Abstract: The selection of an open reading frame (ORF) for translation of eukaryotic mRNA relies on remodeling of the scanning 48S initiation complex into an elongation-ready 80S ribosome. Using cryo-electron ...The selection of an open reading frame (ORF) for translation of eukaryotic mRNA relies on remodeling of the scanning 48S initiation complex into an elongation-ready 80S ribosome. Using cryo-electron microscopy, we visualize the key commitment steps orchestrating 48S remodeling in humans. The mRNA Kozak sequence facilitates mRNA scanning in the 48S open state and stabilizes the 48S closed state by organizing the contacts of eukaryotic initiation factors (eIFs) and ribosomal proteins and by reconfiguring mRNA structure. GTPase-triggered large-scale fluctuations of 48S-bound eIF2 facilitate eIF5B recruitment, transfer of initiator tRNA from eIF2 to eIF5B and the release of eIF5 and eIF2. The 48S-bound multisubunit eIF3 complex controls ribosomal subunit joining by coupling eIF exchange to gradual displacement of the eIF3c N-terminal domain from the intersubunit interface. These findings reveal the structural mechanism of ORF selection in human cells and explain how eIF3 could function in the context of the 80S ribosome. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8rg0.cif.gz | 832.4 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8rg0.ent.gz | 593.6 KB | Display | PDB format |
| PDBx/mmJSON format | 8rg0.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/rg/8rg0ftp://data.pdbj.org/pub/pdb/validation_reports/rg/8rg0 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  19128MC  8pj1C  8pj2C  8pj3C  8pj4C  8pj5C  8pj6C C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj


































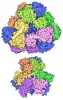



- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Eukaryotic translation initiation factor 3 subunit ... , 9 types, 9 molecules 34568uvxy
| #1: Protein | Mass: 25083.619 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q9UBQ5 |
|---|---|
| #2: Protein | Mass: 37593.645 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: O00303, ubiquitinyl hydrolase 1 |
| #3: Protein | Mass: 66803.734 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q9Y262 |
| #4: Protein | Mass: 42555.832 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q7L2H7 |
| #6: Protein | Mass: 39979.277 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: O15372 |
| #16: Protein | Mass: 166903.781 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q14152 |
| #17: Protein | Mass: 52281.633 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P60228 |
| #18: Protein | Mass: 64060.758 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: O15371 |
| #19: Protein | Mass: 105503.945 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q99613 |
-RNA chain , 2 types, 2 molecules 7A
| #5: RNA chain | Mass: 82412.445 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Production host:  |
|---|---|
| #7: RNA chain | Mass: 603245.500 Da / Num. of mol.: 1 / Source method: isolated from a natural source Details: Only small part of the 18S rRNA is provided with sequence truncated to visualized only parts. Source: (natural) Homo sapiens (human) |
-40S ribosomal protein ... , 8 types, 8 molecules HIMNOPQn
| #8: Protein | Mass: 9480.186 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P42677 |
|---|---|
| #9: Protein | Mass: 17259.389 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62277 |
| #10: Protein | Mass: 15578.156 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P08708 |
| #11: Protein | Mass: 32883.938 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P08865 |
| #12: Protein | Mass: 30002.061 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P61247 |
| #13: Protein | Mass: 16302.772 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62263 |
| #14: Protein | Mass: 13047.532 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62854 |
| #15: Protein | Mass: 7855.052 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62857 |
-Non-polymers , 2 types, 2 molecules 

| #20: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg |
|---|---|
| #21: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn |
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: | ||||||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | ||||||||||||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K / Details: Manual blotting & plunge-freezing |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: SPOT SCAN FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: SPOT SCAN |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 59000 X / Nominal defocus max: 2500 nm / Nominal defocus min: 200 nm / Cs: 0.01 mm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 1.5 sec. / Electron dose: 45 e/Å2 / Detector mode: INTEGRATING / Film or detector model: FEI FALCON III (4k x 4k) |
| EM imaging optics | Spherical aberration corrector: Electron-optical aberrations were corrected using a CETCOR Cs-corrector (CEOS, Heidelberg) aligned with the CETCORPLUS 4.6.9 software package (CEOS, Heidelberg). |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||
| 3D reconstruction | Resolution: 3.4 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 356632 / Symmetry type: POINT | ||||||||||||
| Atomic model building | Space: REAL |
