 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8ott | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | MYC-MAX bound to a nucleosome at SHL+5.8 | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | TRANSCRIPTION / E-box | |||||||||
| Function / homology |  Function and homology information Function and homology informationMad-Max complex / Deposition of new CENPA-containing nucleosomes at the centromere / Inhibition of DNA recombination at telomere / positive regulation of metanephric cap mesenchymal cell proliferation / positive regulation of acinar cell proliferation / acinar cell proliferation / SCF ubiquitin ligase complex binding / NK T cell proliferation / Myc-Max complex / DNA Damage/Telomere Stress Induced Senescence ...Mad-Max complex / Deposition of new CENPA-containing nucleosomes at the centromere / Inhibition of DNA recombination at telomere / positive regulation of metanephric cap mesenchymal cell proliferation / positive regulation of acinar cell proliferation / acinar cell proliferation / SCF ubiquitin ligase complex binding / NK T cell proliferation / Myc-Max complex / DNA Damage/Telomere Stress Induced Senescence / Regulation of endogenous retroelements by KRAB-ZFP proteins / Condensation of Prophase Chromosomes / Negative Regulation of CDH1 Gene Transcription / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / HDACs deacetylate histones / regulation of somatic stem cell population maintenance / regulation of cell cycle process / PRC2 methylates histones and DNA / Metalloprotease DUBs / RNA polymerase II transcription repressor complex / Binding of TCF/LEF:CTNNB1 to target gene promoters / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / cellular response to interferon-alpha / positive regulation of B cell apoptotic process / UCH proteinases / RUNX3 regulates WNT signaling / TFAP2 (AP-2) family regulates transcription of cell cycle factors / myotube differentiation / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / negative regulation of transcription initiation by RNA polymerase II / RMTs methylate histone arginines / negative regulation of cell division / Regulation of CDH1 mRNA translation by microRNAs / negative regulation of monocyte differentiation / detection of mechanical stimulus involved in sensory perception of sound / response to growth factor / B cell apoptotic process / Ub-specific processing proteases / response to alkaloid / transcription regulator activator activity / Transcription of E2F targets under negative control by DREAM complex / fibroblast apoptotic process / negative regulation of stress-activated MAPK cascade / Regulation of NFE2L2 gene expression / protein-DNA complex disassembly / positive regulation of mesenchymal cell proliferation / skeletal system morphogenesis / regulation of telomere maintenance / branching involved in ureteric bud morphogenesis / Signaling by ALK / middle ear morphogenesis / rRNA metabolic process / negative regulation of gene expression via chromosomal CpG island methylation / E-box binding / pigmentation / Transcriptional Regulation by E2F6 / positive regulation of telomere maintenance / MLL1 complex / skeletal muscle cell differentiation / chromosome organization / positive regulation of transcription initiation by RNA polymerase II / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / negative regulation of tumor necrosis factor-mediated signaling pathway / Cyclin E associated events during G1/S transition / core promoter sequence-specific DNA binding / Cyclin A:Cdk2-associated events at S phase entry / negative regulation of fibroblast proliferation / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / ERK1 and ERK2 cascade / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / epigenetic regulation of gene expression / Deposition of new CENPA-containing nucleosomes at the centromere / telomere organization / Interleukin-7 signaling / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / positive regulation of epithelial cell proliferation / Meiotic synapsis / SUMOylation of chromatin organization proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / transcription coregulator binding / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / innate immune response in mucosa Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human)synthetic construct (others) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.3 Å | |||||||||
 Authors Authors | Stoos, L. / Michael, A.K. / Kempf, G. / Kater, L. / Cavadini, S. / Thoma, N. | |||||||||
| Funding support | European Union,  France, 2items France, 2items
| |||||||||
 Citation Citation | Journal: Nature / Year: 2023 Title: Cooperation between bHLH transcription factors and histones for DNA access. Authors: Alicia K Michael / Lisa Stoos / Priya Crosby / Nikolas Eggers / Xinyu Y Nie / Kristina Makasheva / Martina Minnich / Kelly L Healy / Joscha Weiss / Georg Kempf / Simone Cavadini / Lukas ...Authors: Alicia K Michael / Lisa Stoos / Priya Crosby / Nikolas Eggers / Xinyu Y Nie / Kristina Makasheva / Martina Minnich / Kelly L Healy / Joscha Weiss / Georg Kempf / Simone Cavadini / Lukas Kater / Jan Seebacher / Luca Vecchia / Deyasini Chakraborty / Luke Isbel / Ralph S Grand / Florian Andersch / Jennifer L Fribourgh / Dirk Schübeler / Johannes Zuber / Andrew C Liu / Peter B Becker / Beat Fierz / Carrie L Partch / Jerome S Menet / Nicolas H Thomä /     Abstract: The basic helix-loop-helix (bHLH) family of transcription factors recognizes DNA motifs known as E-boxes (CANNTG) and includes 108 members. Here we investigate how chromatinized E-boxes are engaged ...The basic helix-loop-helix (bHLH) family of transcription factors recognizes DNA motifs known as E-boxes (CANNTG) and includes 108 members. Here we investigate how chromatinized E-boxes are engaged by two structurally diverse bHLH proteins: the proto-oncogene MYC-MAX and the circadian transcription factor CLOCK-BMAL1 (refs. ). Both transcription factors bind to E-boxes preferentially near the nucleosomal entry-exit sites. Structural studies with engineered or native nucleosome sequences show that MYC-MAX or CLOCK-BMAL1 triggers the release of DNA from histones to gain access. Atop the H2A-H2B acidic patch, the CLOCK-BMAL1 Per-Arnt-Sim (PAS) dimerization domains engage the histone octamer disc. Binding of tandem E-boxes at endogenous DNA sequences occurs through direct interactions between two CLOCK-BMAL1 protomers and histones and is important for circadian cycling. At internal E-boxes, the MYC-MAX leucine zipper can also interact with histones H2B and H3, and its binding is indirectly enhanced by OCT4 elsewhere on the nucleosome. The nucleosomal E-box position and the type of bHLH dimerization domain jointly determine the histone contact, the affinity and the degree of competition and cooperativity with other nucleosome-bound factors. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8ott.cif.gz | 342 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8ott.ent.gz | 257.5 KB | Display | PDB format |
| PDBx/mmJSON format | 8ott.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ot/8ottftp://data.pdbj.org/pub/pdb/validation_reports/ot/8ott | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  17184MC  8osjC  8oskC  8oslC 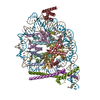 8otsC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |









-Links
 PDBj
PDBj
















































- Assembly
Assembly
| Deposited unit | 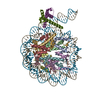
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 8 molecules AEBFDHMN
| #1: Protein | Mass: 11179.090 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: H3C1, H3FA, HIST1H3A, H3C2, H3FL, HIST1H3B, H3C3, H3FC HIST1H3C, H3C4, H3FB, HIST1H3D, H3C6, H3FD, HIST1H3E, H3C7, H3FI, HIST1H3F, H3C8, H3FH, HIST1H3G, H3C10, H3FK, HIST1H3H, H3C11, H3FF, ...Gene: H3C1, H3FA, HIST1H3A, H3C2, H3FL, HIST1H3B, H3C3, H3FC HIST1H3C, H3C4, H3FB, HIST1H3D, H3C6, H3FD, HIST1H3E, H3C7, H3FI, HIST1H3F, H3C8, H3FH, HIST1H3G, H3C10, H3FK, HIST1H3H, H3C11, H3FF, HIST1H3I, H3C12, H3FJ, HIST1H3J Production host:  #2: Protein | Mass: 9279.875 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, ...Gene: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, HIST1H4H, H4C9, H4/M, H4FM, HIST1H4I, H4C11, H4/E, H4FE, HIST1H4J, H4C12, H4/D, H4FD, HIST1H4K, H4C13, H4/K, H4FK, HIST1H4L, H4C14, H4/N, H4F2, H4FN, HIST2H4, HIST2H4A, H4C15, H4/O, H4FO, HIST2H4B, H4C16, H4-16, HIST4H4 Production host: #4: Protein | Mass: 10334.827 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: H2BC11, H2BFR, HIST1H2BJ / Production host: #8: Protein | | Mass: 6346.392 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: MYC, BHLHE39 / Production host: #9: Protein | | Mass: 6031.737 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: MAX, BHLHD4 / Production host: |
|---|
-Histone H2A type 1- ... , 2 types, 2 molecules CG
| #3: Protein | Mass: 11896.886 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: H2AC4, H2AFM, HIST1H2AB, H2AC8, H2AFA, HIST1H2AE / Production host: |
|---|---|
| #5: Protein | Mass: 11807.769 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: H2ac15, Hist1h2ak / Production host: |
-DNA chain , 2 types, 2 molecules IJ
| #6: DNA chain | Mass: 44225.145 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
|---|---|
| #7: DNA chain | Mass: 44674.430 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
-Non-polymers , 1 types, 8 molecules 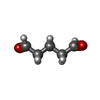
| #10: Chemical | ChemComp-PTD /  Mass: 100.116 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: C5H8O2 Mass: 100.116 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: C5H8O2 |
|---|
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight |
| ||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.2 | ||||||||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 1800 nm / Nominal defocus min: 800 nm |
| Image recording | Electron dose: 50 e/Å2 / Film or detector model: FEI FALCON IV (4k x 4k) |
- Processing
Processing
| CTF correction | Type: NONE |
|---|---|
| 3D reconstruction | Resolution: 3.3 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 152914 / Symmetry type: POINT |
