Synthesis, secretion, and deacylation of Ghrelin / regulation of lipoprotein lipid oxidation / cellular response to L-ascorbic acid / positive regulation of fat cell apoptotic process / negative regulation of glutamine transport / leptin receptor activity / regulation of transport / negative regulation of appetite by leptin-mediated signaling pathway / Synthesis, secretion, and inactivation of Glucagon-like Peptide-1 (GLP-1) / negative regulation of glucagon secretion ...Synthesis, secretion, and deacylation of Ghrelin / regulation of lipoprotein lipid oxidation / cellular response to L-ascorbic acid / positive regulation of fat cell apoptotic process / negative regulation of glutamine transport / leptin receptor activity / regulation of transport / negative regulation of appetite by leptin-mediated signaling pathway / Synthesis, secretion, and inactivation of Glucagon-like Peptide-1 (GLP-1) / negative regulation of glucagon secretion / regulation of endothelial cell proliferation / regulation of natural killer cell mediated cytotoxicity / regulation of natural killer cell proliferation / leptin receptor binding / positive regulation of luteinizing hormone secretion / bone growth / regulation of natural killer cell activation / glycerol biosynthetic process / elastin metabolic process / leptin-mediated signaling pathway / positive regulation of hepatic stellate cell activation / positive regulation of follicle-stimulating hormone secretion / regulation of steroid biosynthetic process / positive regulation of monoatomic ion transport / regulation of intestinal cholesterol absorption / regulation of bone remodeling / regulation of brown fat cell differentiation / positive regulation of peroxisome proliferator activated receptor signaling pathway / regulation of nitric-oxide synthase activity / adult feeding behavior / response to leptin / activation of protein kinase C activity / bone mineralization involved in bone maturation / regulation of lipid biosynthetic process / negative regulation of cartilage development / sexual reproduction / regulation of feeding behavior / ovulation from ovarian follicle / negative regulation of appetite / positive regulation of developmental growth / leukocyte tethering or rolling / energy reserve metabolic process / bile acid metabolic process / cellular response to leptin stimulus / negative regulation of D-glucose import / prostaglandin secretion / regulation of protein localization to nucleus / cardiac muscle hypertrophy / hormone metabolic process / positive regulation of p38MAPK cascade / intestinal absorption / regulation of fat cell differentiation / insulin secretion / regulation of metabolic process / aorta development / negative regulation of vasoconstriction / regulation of gluconeogenesis / peptide hormone receptor binding / glycogen metabolic process / eating behavior / fatty acid beta-oxidation / regulation of cytokine production involved in inflammatory response / central nervous system neuron development / response to dietary excess / negative regulation of lipid storage / T cell differentiation / response to vitamin E / cell surface receptor signaling pathway via JAK-STAT / positive regulation of TOR signaling / regulation of angiogenesis / adipose tissue development / negative regulation of gluconeogenesis / phagocytosis / positive regulation of insulin receptor signaling pathway / glial cell proliferation / positive regulation of T cell proliferation / energy homeostasis / cellular response to retinoic acid / positive regulation of interleukin-12 production / regulation of insulin secretion / negative regulation of autophagy / cholesterol metabolic process / response to activity / gluconeogenesis / positive regulation of interleukin-8 production / female pregnancy / positive regulation of receptor signaling pathway via JAK-STAT / determination of adult lifespan / response to insulin / placenta development / hormone activity / regulation of blood pressure / lipid metabolic process / positive regulation of interleukin-6 production / positive regulation of protein import into nucleus / cellular response to insulin stimulus / circadian rhythm / glucose metabolic process / positive regulation of reactive oxygen species metabolic process / positive regulation of tumor necrosis factor production 類似検索 - 分子機能
Leptin / Leptin / Leptin receptor, immunoglobulin-like domain / Obesity receptor immunoglobulin like domain / Short hematopoietin receptor, family 1, conserved site / Immunoglobulin C2-set-like, ligand-binding / Ig-like C2-type domain / Long hematopoietin receptor, Gp130 family 2, conserved site / Long hematopoietin receptor, gp130 family signature. / Growth Hormone; Chain: A; - #10 ...Leptin / Leptin / Leptin receptor, immunoglobulin-like domain / Obesity receptor immunoglobulin like domain / Short hematopoietin receptor, family 1, conserved site / Immunoglobulin C2-set-like, ligand-binding / Ig-like C2-type domain / Long hematopoietin receptor, Gp130 family 2, conserved site / Long hematopoietin receptor, gp130 family signature. / Growth Hormone; Chain: A; - #10 / Four-helical cytokine-like, core / Growth Hormone; Chain: A; / Fibronectin type III domain / Fibronectin type 3 domain / Fibronectin type-III domain profile. / Fibronectin type III / Fibronectin type III superfamily / Ig-like domain profile. / Immunoglobulin-like domain / Immunoglobulin-like fold / Up-down Bundle / Mainly Alpha 類似検索 - ドメイン・相同性
ジャーナル: Nat Struct Mol Biol / 年: 2023 タイトル: Mechanism of receptor assembly via the pleiotropic adipokine Leptin. 著者: Alexandra Tsirigotaki / Ann Dansercoer / Koen H G Verschueren / Iva Marković / Christoph Pollmann / Maximillian Hafer / Jan Felix / Catherine Birck / Wouter Van Putte / Dominiek Catteeuw / ...著者: Alexandra Tsirigotaki / Ann Dansercoer / Koen H G Verschueren / Iva Marković / Christoph Pollmann / Maximillian Hafer / Jan Felix / Catherine Birck / Wouter Van Putte / Dominiek Catteeuw / Jan Tavernier / J Fernando Bazan / Jacob Piehler / Savvas N Savvides / Kenneth Verstraete / 要旨: The adipokine Leptin activates its receptor LEP-R in the hypothalamus to regulate body weight and exerts additional pleiotropic functions in immunity, fertility and cancer. However, the structure and ...The adipokine Leptin activates its receptor LEP-R in the hypothalamus to regulate body weight and exerts additional pleiotropic functions in immunity, fertility and cancer. However, the structure and mechanism of Leptin-mediated LEP-R assemblies has remained unclear. Intriguingly, the signaling-competent isoform of LEP-R is only lowly abundant amid several inactive short LEP-R isoforms contributing to a mechanistic conundrum. Here we show by X-ray crystallography and cryo-EM that, in contrast to long-standing paradigms, Leptin induces type I cytokine receptor assemblies featuring 3:3 stoichiometry and demonstrate such Leptin-induced trimerization of LEP-R on living cells via single-molecule microscopy. In mediating these assemblies, Leptin undergoes drastic restructuring that activates its site III for binding to the Ig domain of an adjacent LEP-R. These interactions are abolished by mutations linked to obesity. Collectively, our study provides the structural and mechanistic framework for how evolutionarily conserved Leptin:LEP-R assemblies with 3:3 stoichiometry can engage distinct LEP-R isoforms to achieve signaling.
分子量: 16220.559 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: Mouse leptin (Uniprot ID P41160) was refolded from inclusion bodies produced in E.coli. 由来: (組換発現) Mus musculus (ハツカネズミ) / 遺伝子: Lep, Ob / プラスミド: pET11d / 発現宿主: Escherichia coli BL21(DE3) (大腸菌) / 参照: UniProt: P41160
#2: タンパク質
Leptinreceptor / LEP-R / B219 / OB receptor / OB-R
分子量: 37429.477 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: The mouse LepR-IgCRH2 fragment was produced in HEK293 Freestyle cells in the presence of kifunensine. Prior to crystallisation, the N-terminal His-tag was removed with TEV protease and the N- ...詳細: The mouse LepR-IgCRH2 fragment was produced in HEK293 Freestyle cells in the presence of kifunensine. Prior to crystallisation, the N-terminal His-tag was removed with TEV protease and the N-terminal glycans were trimmed with EndoH. 由来: (組換発現) Mus musculus (ハツカネズミ) / 遺伝子: Lepr, Db, Obr / プラスミド: pTwist CMV BetaGlobin / 発現宿主: Homo sapiens (ヒト) / 株 (発現宿主): FreeStyle / 参照: UniProt: P48356
解像度: 2.951→66.34 Å / Cor.coef. Fo:Fc: 0.943 / Cor.coef. Fo:Fc free: 0.927 / SU R Cruickshank DPI: 0.633 / 交差検証法: THROUGHOUT / SU R Blow DPI: 0.597 / SU Rfree Blow DPI: 0.32 / SU Rfree Cruickshank DPI: 0.328
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 X線回折 /
X線回折 /  データ登録者
データ登録者 ベルギー, 1件
ベルギー, 1件  引用
引用


 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード

















 PDBj
PDBj
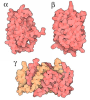


 集合体
集合体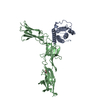


 Homo sapiens (ヒト) / 株 (発現宿主): FreeStyle / 参照: UniProt: P48356
Homo sapiens (ヒト) / 株 (発現宿主): FreeStyle / 参照: UniProt: P48356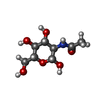
 タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C8H15NO6
タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C8H15NO6 分子量: 18.015 Da / 分子数: 7 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 7 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析