| 登録情報 | データベース: PDB / ID: 7okp
|
|---|
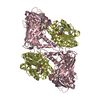
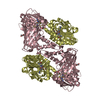
| タイトル | Crystal structure of mouse CARM1 in complex with histone H3_13-22 K18 acetylated |
|---|
 要素 要素 | - Histone H3.3
- Histone-arginine methyltransferase CARM1
|
|---|
 キーワード キーワード | TRANSCRIPTION / protein arginine N-methyltransferase / PRMT / CARM1 / transition state mimics |
|---|
| 機能・相同性 |  機能・相同性情報 機能・相同性情報
regulation of growth plate cartilage chondrocyte proliferation / histone H3R26 methyltransferase activity / histone H3R17 methyltransferase activity / endochondral bone morphogenesis / histone H3R2 methyltransferase activity / RMTs methylate histone arginines / Regulation of lipid metabolism by PPARalpha / negative regulation of chromosome condensation / Cytoprotection by HMOX1 / protein-arginine omega-N asymmetric methyltransferase activity ...regulation of growth plate cartilage chondrocyte proliferation / histone H3R26 methyltransferase activity / histone H3R17 methyltransferase activity / endochondral bone morphogenesis / histone H3R2 methyltransferase activity / RMTs methylate histone arginines / Regulation of lipid metabolism by PPARalpha / negative regulation of chromosome condensation / Cytoprotection by HMOX1 / protein-arginine omega-N asymmetric methyltransferase activity / Barr body / type I protein arginine methyltransferase / : / Estrogen-dependent gene expression / protein methyltransferase activity / regulation of intracellular estrogen receptor signaling pathway / replication fork reversal / inner kinetochore / pericentric heterochromatin formation / protein-arginine N-methyltransferase activity / muscle cell differentiation / oocyte maturation / nucleosomal DNA binding / histone methyltransferase activity / nucleus organization / nuclear replication fork / spermatid development / positive regulation of fat cell differentiation / nuclear receptor-mediated steroid hormone signaling pathway / single fertilization / subtelomeric heterochromatin formation / RNA polymerase II core promoter sequence-specific DNA binding / Replacement of protamines by nucleosomes in the male pronucleus / estrogen receptor signaling pathway / protein localization to chromatin / embryo implantation / telomere organization / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / NoRC negatively regulates rRNA expression / B-WICH complex positively regulates rRNA expression / Meiotic recombination / Pre-NOTCH Transcription and Translation / RNA polymerase II transcription regulator complex / Activation of anterior HOX genes in hindbrain development during early embryogenesis / multicellular organism growth / male gonad development / Transcriptional regulation of granulopoiesis / osteoblast differentiation / structural constituent of chromatin / nucleosome / RUNX1 regulates transcription of genes involved in differentiation of HSCs / nucleosome assembly / Senescence-Associated Secretory Phenotype (SASP) / Factors involved in megakaryocyte development and platelet production / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / positive regulation of cell growth / Oxidative Stress Induced Senescence / methylation / Estrogen-dependent gene expression / transcription coactivator activity / chromosome, telomeric region / cell population proliferation / transcription cis-regulatory region binding / RNA polymerase II cis-regulatory region sequence-specific DNA binding / Amyloid fiber formation / protein heterodimerization activity / positive regulation of cell population proliferation / regulation of DNA-templated transcription / positive regulation of transcription by RNA polymerase II / protein-containing complex / extracellular exosome / extracellular region / nucleoplasm / nucleus / cytosol類似検索 - 分子機能 Histone-arginine methyltransferase CARM1, N-terminal / Coactivator-associated arginine methyltransferase 1 N terminal / Ribosomal protein L11 methyltransferase (PrmA) / Hnrnp arginine n-methyltransferase1 / Hnrnp arginine n-methyltransferase1 / : / Arginine methyltransferase oligomerization subdomain / Protein arginine N-methyltransferase / SAM-dependent methyltransferase PRMT-type domain profile. / Vaccinia Virus protein VP39 ...Histone-arginine methyltransferase CARM1, N-terminal / Coactivator-associated arginine methyltransferase 1 N terminal / Ribosomal protein L11 methyltransferase (PrmA) / Hnrnp arginine n-methyltransferase1 / Hnrnp arginine n-methyltransferase1 / : / Arginine methyltransferase oligomerization subdomain / Protein arginine N-methyltransferase / SAM-dependent methyltransferase PRMT-type domain profile. / Vaccinia Virus protein VP39 / Distorted Sandwich / Histone H3 signature 1. / Histone H3 signature 2. / Histone H3 / Histone H3/CENP-A / Histone H2A/H2B/H3 / Core histone H2A/H2B/H3/H4 / Histone-fold / PH-like domain superfamily / S-adenosyl-L-methionine-dependent methyltransferase superfamily / Rossmann fold / 3-Layer(aba) Sandwich / Mainly Beta / Alpha Beta類似検索 - ドメイン・相同性 MALONATE ION / Chem-QVR / Histone H3.3 / Histone-arginine methyltransferase CARM1類似検索 - 構成要素 |
|---|
| 生物種 |   Mus musculus (ハツカネズミ) Mus musculus (ハツカネズミ)
 Homo sapiens (ヒト) Homo sapiens (ヒト) |
|---|
| 手法 |  X線回折 / 分子置換 / 解像度: 2.2 Å X線回折 / 分子置換 / 解像度: 2.2 Å |
|---|
 データ登録者 データ登録者 | Marechal, N. / Cura, V. / Troffer-Charlier, N. / Bonnefond, L. / Cavarelli, J. |
|---|
 引用 引用 | ジャーナル: Chembiochem / 年: 2021
タイトル: Structural Studies Provide New Insights into the Role of Lysine Acetylation on Substrate Recognition by CARM1 and Inform the Design of Potent Peptidomimetic Inhibitors.
著者: Zhang, Y. / Marechal, N. / van Haren, M.J. / Troffer-Charlier, N. / Cura, V. / Cavarelli, J. / Martin, N.I. |
|---|
| 履歴 | | 登録 | 2021年5月18日 | 登録サイト: PDBE / 処理サイト: PDBE |
|---|
| 改定 1.0 | 2021年10月27日 | Provider: repository / タイプ: Initial release |
|---|
| 改定 1.1 | 2021年12月22日 | Group: Database references / カテゴリ: citation
Item: _citation.journal_volume / _citation.page_first / _citation.page_last |
|---|
| 改定 2.0 | 2023年11月15日 | Group: Atomic model / Data collection / カテゴリ: atom_site / chem_comp_atom / chem_comp_bond / Item: _atom_site.auth_atom_id / _atom_site.label_atom_id |
|---|
| 改定 2.1 | 2024年1月31日 | Group: Refinement description / カテゴリ: pdbx_initial_refinement_model |
|---|
| 改定 2.2 | 2024年11月6日 | Group: Structure summary
カテゴリ: pdbx_entry_details / pdbx_modification_feature
Item: _pdbx_entry_details.has_protein_modification |
|---|
|
|---|
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード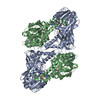




 PDBj
PDBj










 集合体
集合体


 Spodoptera aff. frugiperda 1 BOLD-2017 (蝶・蛾)
Spodoptera aff. frugiperda 1 BOLD-2017 (蝶・蛾)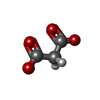
 分子量: 102.046 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H2O4
分子量: 102.046 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H2O4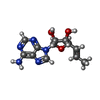
 分子量: 277.279 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C12H15N5O3 / タイプ: SUBJECT OF INVESTIGATION
分子量: 277.279 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C12H15N5O3 / タイプ: SUBJECT OF INVESTIGATION 分子量: 18.015 Da / 分子数: 421 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 421 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析