ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7o3c | ||||||
|---|---|---|---|---|---|---|---|
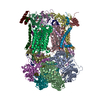
| タイトル | Murine supercomplex CIII2CIV in the mature unlocked conformation | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | MEMBRANE PROTEIN / mitochondria / respiratory chain / supercomplex / OXIDOREDUCTASE / complex III / complex IV | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Complex IV assembly / response to D-galactosamine / Complex III assembly / TP53 Regulates Metabolic Genes / regulation of oxidative phosphorylation / respiratory chain complex IV assembly / response to mercury ion / subthalamus development / pons development / Cytoprotection by HMOX1 ...Complex IV assembly / response to D-galactosamine / Complex III assembly / TP53 Regulates Metabolic Genes / regulation of oxidative phosphorylation / respiratory chain complex IV assembly / response to mercury ion / subthalamus development / pons development / Cytoprotection by HMOX1 / Respiratory electron transport / mitochondrial respirasome assembly / response to cobalamin / cerebellar Purkinje cell layer development / aerobic electron transport chain / mitochondrial respiratory chain complex III assembly / pyramidal neuron development / response to alkaloid / respiratory chain complex IV / cellular respiration / thalamus development / Mitochondrial protein degradation / respiratory chain complex / cytochrome-c oxidase / respiratory chain complex III / oxidative phosphorylation / quinol-cytochrome-c reductase / mitochondrial electron transport, cytochrome c to oxygen / response to glucagon / quinol-cytochrome-c reductase activity / cytochrome-c oxidase activity / response to copper ion / mitochondrial electron transport, ubiquinol to cytochrome c / hypothalamus development / midbrain development / electron transport coupled proton transport / response to electrical stimulus / animal organ regeneration / response to hyperoxia / response to cadmium ion / response to hormone / lactation / response to nutrient / cerebellum development / central nervous system development / response to activity / respiratory electron transport chain / hippocampus development / electron transport chain / response to calcium ion / mitochondrial membrane / metalloendopeptidase activity / 2 iron, 2 sulfur cluster binding / response to toxic substance / myelin sheath / response to oxidative stress / response to ethanol / protein-macromolecule adaptor activity / response to hypoxia / oxidoreductase activity / mitochondrial inner membrane / response to xenobiotic stimulus / copper ion binding / heme binding / ubiquitin protein ligase binding / protein-containing complex binding / mitochondrion / proteolysis / nucleoplasm / metal ion binding / membrane / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.3 Å | ||||||
 データ登録者 データ登録者 | Vercellino, I. / Sazanov, L.A. | ||||||
| 資金援助 |  オーストリア, 1件 オーストリア, 1件
| ||||||
 引用 引用 | ジャーナル: Nature / 年: 2021 タイトル: Structure and assembly of the mammalian mitochondrial supercomplex CIIICIV. 著者: Irene Vercellino / Leonid A Sazanov / 要旨: The enzymes of the mitochondrial electron transport chain are key players of cell metabolism. Despite being active when isolated, in vivo they associate into supercomplexes, whose precise role is ...The enzymes of the mitochondrial electron transport chain are key players of cell metabolism. Despite being active when isolated, in vivo they associate into supercomplexes, whose precise role is debated. Supercomplexes CIIICIV (refs. ), CICIII (ref. ) and CICIIICIV (respirasome) exist in mammals, but in contrast to CICIII and the respirasome, to date the only known eukaryotic structures of CIIICIV come from Saccharomyces cerevisiae and plants, which have different organization. Here we present the first, to our knowledge, structures of mammalian (mouse and ovine) CIIICIV and its assembly intermediates, in different conformations. We describe the assembly of CIIICIV from the CIII precursor to the final CIIICIV conformation, driven by the insertion of the N terminus of the assembly factor SCAF1 (ref. ) deep into CIII, while its C terminus is integrated into CIV. Our structures (which include CICIII and the respirasome) also confirm that SCAF1 is exclusively required for the assembly of CIIICIV and has no role in the assembly of the respirasome. We show that CIII is asymmetric due to the presence of only one copy of subunit 9, which straddles both monomers and prevents the attachment of a second copy of SCAF1 to CIII, explaining the presence of one copy of CIV in CIIICIV in mammals. Finally, we show that CIII and CIV gain catalytic advantage when assembled into the supercomplex and propose a role for CIIICIV in fine tuning the efficiency of electron transfer in the electron transport chain. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7o3c.cif.gz | 1.4 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7o3c.ent.gz | 992.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7o3c.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7o3c_validation.pdf.gz | 2.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7o3c_full_validation.pdf.gz | 2.7 MB | 表示 | |
| XML形式データ | 7o3c_validation.xml.gz | 153.4 KB | 表示 | |
| CIF形式データ | 7o3c_validation.cif.gz | 229.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/o3/7o3cftp://data.pdbj.org/pub/pdb/validation_reports/o3/7o3c | HTTPS FTP |
-関連構造データ








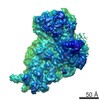
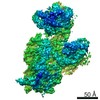
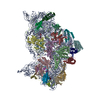
-リンク
 PDBj
PDBj

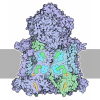





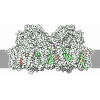

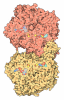

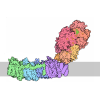




- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-Cytochrome b-c1 complex subunit ... , 9種, 17分子 ALBMEPFQGRHSJUKVT
| #1: タンパク質 | 分子量: 49355.301 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #2: タンパク質 | 分子量: 46641.773 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #5: タンパク質 | 分子量: 21524.398 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #6: タンパク質 | 分子量: 13456.336 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #7: タンパク質 | 分子量: 9652.051 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #8: タンパク質 | 分子量: 9002.015 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #9: タンパク質 | 分子量: 7326.330 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #10: タンパク質 | 分子量: 6546.627 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #11: タンパク質 | | 分子量: 7900.234 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-タンパク質 , 3種, 5分子 CNDOI
| #3: タンパク質 | 分子量: 43240.305 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #4: タンパク質 | 分子量: 27312.188 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #12: タンパク質 | | 分子量: 12575.581 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-Cytochrome c oxidase subunit ... , 12種, 12分子 abcdefghiklm
| #13: タンパク質 | 分子量: 56945.641 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #14: タンパク質 | 分子量: 25993.318 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #15: タンパク質 | 分子量: 29948.689 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #16: タンパク質 | 分子量: 17225.596 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #17: タンパク質 | 分子量: 12453.081 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #18: タンパク質 | 分子量: 10862.376 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #19: タンパク質 | 分子量: 9437.691 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #20: タンパク質 | 分子量: 9955.270 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #21: タンパク質 | 分子量: 8352.825 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #22: タンパク質 | 分子量: 6317.091 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #23: タンパク質・ペプチド | 分子量: 5451.369 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #24: タンパク質・ペプチド | 分子量: 4660.410 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-非ポリマー , 13種, 41分子 








| #25: 化合物 | ChemComp-3PE /  分子量: 748.065 Da / 分子数: 15 / 由来タイプ: 合成 / 式: C41H82NO8P / コメント: リン脂質*YM 分子量: 748.065 Da / 分子数: 15 / 由来タイプ: 合成 / 式: C41H82NO8P / コメント: リン脂質*YM#26: 化合物 | ChemComp-CDL /  分子量: 1464.043 Da / 分子数: 7 / 由来タイプ: 合成 / 式: C81H156O17P2 / コメント: リン脂質*YM 分子量: 1464.043 Da / 分子数: 7 / 由来タイプ: 合成 / 式: C81H156O17P2 / コメント: リン脂質*YM#27: 化合物 | ChemComp-HEM /  分子量: 616.487 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C34H32FeN4O4 分子量: 616.487 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C34H32FeN4O4#28: 化合物 |  分子量: 618.503 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C34H34FeN4O4 分子量: 618.503 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C34H34FeN4O4#29: 化合物 |  分子量: 175.820 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Fe2S2 分子量: 175.820 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Fe2S2#30: 化合物 |  分子量: 790.145 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C44H88NO8P / コメント: リン脂質*YM 分子量: 790.145 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C44H88NO8P / コメント: リン脂質*YM#31: 化合物 | ChemComp-CU / |  分子量: 63.546 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Cu 分子量: 63.546 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Cu#32: 化合物 | ChemComp-NA / |  分子量: 22.990 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Na 分子量: 22.990 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Na#33: 化合物 |  分子量: 852.837 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C49H56FeN4O6 分子量: 852.837 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C49H56FeN4O6#34: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg#35: 化合物 | ChemComp-CUA / |  分子量: 127.092 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Cu2 分子量: 127.092 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Cu2#36: 化合物 | ChemComp-ZN / |  分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn#37: 化合物 | ChemComp-TGL / |  分子量: 891.480 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C57H110O6 分子量: 891.480 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C57H110O6 |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Murine full supercomplex CIII2CIV in the unlocked conformation タイプ: COMPLEX 詳細: Dimer of Complex III, monomer of complex IV, bridged by SCAF1 Entity ID: #1-#24 / 由来: NATURAL | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.7 MDa / 実験値: YES | |||||||||||||||||||||||||
| 由来(天然) | 生物種: | |||||||||||||||||||||||||
| 緩衝液 | pH: 7.7 / 詳細: CHAPS was added only upon freezing | |||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||
| 試料 | 濃度: 4 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R0.6/1 | |||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 75000 X / 最大 デフォーカス(公称値): 1600 nm / 最小 デフォーカス(公称値): 400 nm / Calibrated defocus min: 200 nm / 最大 デフォーカス(補正後): 2700 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.17 sec. / 電子線照射量: 90.66 e/Å2 / 検出モード: INTEGRATING フィルム・検出器のモデル: FEI FALCON III (4k x 4k) 撮影したグリッド数: 1 / 実像数: 7245 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1500000 詳細: multiple rounds of picking and classification performed, first with Relion LoG and then with Topaz, to extract the best particles from the non-pure starting material | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 16228 / アルゴリズム: FOURIER SPACE / クラス平均像の数: 1 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 80 / プロトコル: OTHER / 空間: REAL / Target criteria: MAXIMAL LIKELYHOOD | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 80.34 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|