 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6yt3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of the MoStoNano fusion protein | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | STRUCTURAL PROTEIN / Fusion protein / crystal engineering / rigid helix / molecular biomimetics | |||||||||
| Function / homology |  Function and homology information Function and homology informationnutrient reservoir activity / molybdenum ion binding / protein-disulfide reductase activity / cell redox homeostasis / cytosol / cytoplasm Similarity search - Function | |||||||||
| Biological species |  Azotobacter vinelandii (bacteria)Salmonella enterica subsp. enterica serovar Bovismorbificans (bacteria) Azotobacter vinelandii (bacteria)Salmonella enterica subsp. enterica serovar Bovismorbificans (bacteria) | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.85 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.85 Å | |||||||||
 Authors Authors | Benoit, R.M. / Bierig, T. / Collu, C. / Engilberge, S. / Olieric, V. | |||||||||
| Funding support |  Switzerland, 2items Switzerland, 2items
| |||||||||
 Citation Citation | Journal: Structure / Year: 2022 Title: Chimeric single α-helical domains as rigid fusion protein connections for protein nanotechnology and structural biology. Authors: Gabriella Collu / Tobias Bierig / Anna-Sophia Krebs / Sylvain Engilberge / Niveditha Varma / Ramon Guixà-González / Timothy Sharpe / Xavier Deupi / Vincent Olieric / Emiliya Poghosyan / Roger M Benoit / Abstract: Chimeric fusion proteins are essential tools for protein nanotechnology. Non-optimized protein-protein connections are usually flexible and therefore unsuitable as structural building blocks. Here we ...Chimeric fusion proteins are essential tools for protein nanotechnology. Non-optimized protein-protein connections are usually flexible and therefore unsuitable as structural building blocks. Here we show that the ER/K motif, a single α-helical domain (SAH), can be seamlessly fused to terminal helices of proteins, forming an extended, partially free-standing rigid helix. This enables the connection of two domains at a defined distance and orientation. We designed three constructs termed YFPnano, T4Lnano, and MoStoNano. Analysis of experimentally determined structures and molecular dynamics simulations reveals a certain degree of plasticity in the connections that allows the adaptation to crystal contact opportunities. Our data show that SAHs can be stably integrated into designed structural elements, enabling new possibilities for protein nanotechnology, for example, to improve the exposure of epitopes on nanoparticles (structural vaccinology), to engineer crystal contacts with minimal impact on construct flexibility (for the study of protein dynamics), and to design novel biomaterials. #1: Journal: Biorxiv / Year: 2020Title: Chimeric single alpha-helical domains as rigid fusion protein connections for protein nanotechnology and structural biology Authors: Collu, G. / Bierig, T. / Krebs, A.-S. / Engilberge, S. / Varma, N. / Guixa-Gonzalez, R. / Deupi, X. / Olieric, V. / Poghosyan, E. / Benoit, R.M. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6yt3.cif.gz | 260.2 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6yt3.ent.gz | 208.2 KB | Display | PDB format |
| PDBx/mmJSON format | 6yt3.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/yt/6yt3ftp://data.pdbj.org/pub/pdb/validation_reports/yt/6yt3 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  6hr1C  6xyrC  4f6tS C: citing same article ( S: Starting model for refinement |
|---|---|
| Similar structure data |

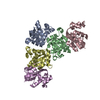
-Links
 PDBj
PDBj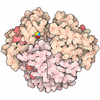

- Assembly
Assembly
| Deposited unit | 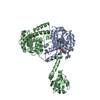
| ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||
| Unit cell |
| ||||||||||||||||||
| Components on special symmetry positions |
|
-Components
| #1: Protein | Mass: 31428.973 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Azotobacter vinelandii (strain DJ / ATCC BAA-1303) (bacteria)Strain: DJ / ATCC BAA-1303 / Gene: mosA, Avin_43200 / Production host: | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 40436.559 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: chain B has three components that are fused together: Thioredoxin at the N-terminus, then a short EEEKRKREEE rigid helix, then the beta-subunit of MoSto. Source: (gene. exp.) Salmonella enterica subsp. enterica serovar Bovismorbificans (bacteria), (gene. exp.) Azotobacter vinelandii (strain DJ / ATCC BAA-1303) (bacteria)Gene: trxA_2, trxA, A3789_21210, AL561_14185, B7N00_20260, B7N01_17730, B7N34_19660, B7N35_19470, B7N60_21445, B7N72_16575, B7N73_22720, B7N78_20580, B7N79_19980, B7N80_16005, B7N84_20570, B7N95_ ...Gene: trxA_2, trxA, A3789_21210, AL561_14185, B7N00_20260, B7N01_17730, B7N34_19660, B7N35_19470, B7N60_21445, B7N72_16575, B7N73_22720, B7N78_20580, B7N79_19980, B7N80_16005, B7N84_20570, B7N95_20865, CAD68_20095, CBK57_22150, DK062_18215, DOI63_23750, DP779_24195, DPA33_18890, DPK57_04940, DPS13_18845, DPZ52_04610, DRU31_19690, E0916_20730, EJV93_21005, ERS008198_02131, ERS008202_02904, ERS008207_01732, EWC73_19305, EXP31_20825, NCTC5754_04586, mosB, Avin_43210 Strain: DJ / ATCC BAA-1303 / Production host: | ||||||||
| #3: Chemical |   Mass: 507.181 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM#4: Chemical | ChemComp-MG / |   Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg#5: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 203 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 203 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | Has protein modification | Y | |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal grow | Temperature: 293 K / Method: vapor diffusion Details: 0.1 M tri-Sodium citrate pH 5.6, 10% PEG 4000, 10% Isopropanol |
|---|
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SLS / Beamline: X06DA / Wavelength: 1 Å |
| Detector | Type: DECTRIS PILATUS 2M / Detector: PIXEL / Date: May 16, 2019 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 2.85→47.89 Å / Num. obs: 33352 / % possible obs: 99.9 % / Redundancy: 10.6 % / CC1/2: 0.998 / Rpim(I) all: 0.071 / Net I/σ(I): 8.2 |
| Reflection shell | Resolution: 2.85→2.98 Å / Mean I/σ(I) obs: 2.3 / Num. unique obs: 1670 / CC1/2: 0.8 / Rpim(I) all: 0.351 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 4F6T Resolution: 2.85→47.89 Å / Cor.coef. Fo:Fc: 0.871 / Cor.coef. Fo:Fc free: 0.851 / SU R Cruickshank DPI: 0.436 / Cross valid method: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.44 / SU Rfree Blow DPI: 0.312 / SU Rfree Cruickshank DPI: 0.315
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 160.57 Å2 / Biso mean: 65.14 Å2 / Biso min: 3 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.47 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: final / Resolution: 2.85→47.89 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 2.85→2.9 Å / Rfactor Rfree error: 0 / Total num. of bins used: 50
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
