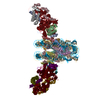
Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6t9l | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | SAGA DUB module bound to a ubiqitinated nucleosome | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | GENE REGULATION / Coactivator / Transcription / Histone acetyltransferase / Histone deubiquitinase | |||||||||
| Function / homology |  Function and homology information Function and homology informationDUBm complex / RITS complex assembly / regulation of nucleocytoplasmic transport / transcription export complex 2 / nuclear mRNA surveillance / regulatory ncRNA-mediated heterochromatin formation / SLIK (SAGA-like) complex / post-transcriptional tethering of RNA polymerase II gene DNA at nuclear periphery / SAGA complex / regulation of protein localization to chromatin ...DUBm complex / RITS complex assembly / regulation of nucleocytoplasmic transport / transcription export complex 2 / nuclear mRNA surveillance / regulatory ncRNA-mediated heterochromatin formation / SLIK (SAGA-like) complex / post-transcriptional tethering of RNA polymerase II gene DNA at nuclear periphery / SAGA complex / regulation of protein localization to chromatin / poly(A)+ mRNA export from nucleus / nuclear pore / positive regulation of RNA polymerase II transcription preinitiation complex assembly / protein deubiquitination / mRNA export from nucleus / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / RNA splicing / FLT3 signaling by CBL mutants / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Prevention of phagosomal-lysosomal fusion / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / Negative regulation of FLT3 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Downregulation of ERBB4 signaling / APC-Cdc20 mediated degradation of Nek2A / Regulation of FZD by ubiquitination / p75NTR recruits signalling complexes / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / NF-kB is activated and signals survival / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / Pexophagy / Downregulation of ERBB2:ERBB3 signaling / NRIF signals cell death from the nucleus / Regulation of PTEN localization / Regulation of innate immune responses to cytosolic DNA / VLDLR internalisation and degradation / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by REV1 / TICAM1, RIP1-mediated IKK complex recruitment / Regulation of BACH1 activity / Translesion synthesis by POLK / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / InlB-mediated entry of Listeria monocytogenes into host cell / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Translesion synthesis by POLI / Downregulation of TGF-beta receptor signaling / Josephin domain DUBs / Gap-filling DNA repair synthesis and ligation in GG-NER / IKK complex recruitment mediated by RIP1 / PINK1-PRKN Mediated Mitophagy / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / TNFR1-induced NF-kappa-B signaling pathway / Regulation of activated PAK-2p34 by proteasome mediated degradation / TCF dependent signaling in response to WNT / Regulation of NF-kappa B signaling / activated TAK1 mediates p38 MAPK activation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / NOTCH3 Activation and Transmission of Signal to the Nucleus / Regulation of signaling by CBL / Negative regulators of DDX58/IFIH1 signaling / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Asymmetric localization of PCP proteins / Negative regulation of FGFR3 signaling / Ubiquitin-dependent degradation of Cyclin D / Fanconi Anemia Pathway / Deactivation of the beta-catenin transactivating complex / Peroxisomal protein import / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / Stabilization of p53 / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / Negative regulation of FGFR2 signaling / Negative regulation of FGFR4 signaling / Downregulation of SMAD2/3:SMAD4 transcriptional activity / Negative regulation of FGFR1 signaling / Termination of translesion DNA synthesis / Assembly of the pre-replicative complex / Vpu mediated degradation of CD4 Similarity search - Function | |||||||||
| Biological species |   Homo sapiens (human) Homo sapiens (human)synthetic construct (others) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.6 Å | |||||||||
 Authors Authors | Wang, H. / Cramer, P. | |||||||||
| Funding support |  Germany, 2items Germany, 2items
| |||||||||
 Citation Citation | Journal: Nature / Year: 2020 Title: Structure of the transcription coactivator SAGA. Authors: Haibo Wang / Christian Dienemann / Alexandra Stützer / Henning Urlaub / Alan C M Cheung / Patrick Cramer /  Abstract: Gene transcription by RNA polymerase II is regulated by activator proteins that recruit the coactivator complexes SAGA (Spt-Ada-Gcn5-acetyltransferase) and transcription factor IID (TFIID). SAGA is ...Gene transcription by RNA polymerase II is regulated by activator proteins that recruit the coactivator complexes SAGA (Spt-Ada-Gcn5-acetyltransferase) and transcription factor IID (TFIID). SAGA is required for all regulated transcription and is conserved among eukaryotes. SAGA contains four modules: the activator-binding Tra1 module, the core module, the histone acetyltransferase (HAT) module and the histone deubiquitination (DUB) module. Previous studies provided partial structures, but the structure of the central core module is unknown. Here we present the cryo-electron microscopy structure of SAGA from the yeast Saccharomyces cerevisiae and resolve the core module at 3.3 Å resolution. The core module consists of subunits Taf5, Sgf73 and Spt20, and a histone octamer-like fold. The octamer-like fold comprises the heterodimers Taf6-Taf9, Taf10-Spt7 and Taf12-Ada1, and two histone-fold domains in Spt3. Spt3 and the adjacent subunit Spt8 interact with the TATA box-binding protein (TBP). The octamer-like fold and its TBP-interacting region are similar in TFIID, whereas Taf5 and the Taf6 HEAT domain adopt distinct conformations. Taf12 and Spt20 form flexible connections to the Tra1 module, whereas Sgf73 tethers the DUB module. Binding of a nucleosome to SAGA displaces the HAT and DUB modules from the core-module surface, allowing the DUB module to bind one face of an ubiquitinated nucleosome. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6t9l.cif.gz | 432 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6t9l.ent.gz | 322 KB | Display | PDB format |
| PDBx/mmJSON format | 6t9l.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/t9/6t9lftp://data.pdbj.org/pub/pdb/validation_reports/t9/6t9l | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  10415MC  6t9iC  6t9jC  6t9kC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj


















































- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 8 types, 11 molecules AEBFCGDHKLO
| #1: Protein | Mass: 15303.930 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  #2: Protein | Mass: 11263.231 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #3: Protein | Mass: 13978.241 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #4: Protein | | Mass: 13524.752 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #5: Protein | | Mass: 13848.097 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #8: Protein | | Mass: 53692.270 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: This is a zinc containing protein. Source: (gene. exp.) Gene: UBP8, YMR223W, YM9959.05 / Production host: #9: Protein | | Mass: 11094.497 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: Q6WNK7 #12: Protein | | Mass: 8576.831 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: UBC / Production host: |
|---|
-Widom601 DNA (145- ... , 2 types, 2 molecules IJ
| #6: DNA chain | Mass: 44520.383 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
|---|---|
| #7: DNA chain | Mass: 44991.660 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
-SAGA-associated factor ... , 2 types, 2 molecules MN
| #10: Protein | Mass: 11297.625 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Details: This a zinc-finger containing protein. Source: (natural) References: UniProt: Q03067 |
|---|---|
| #11: Protein | Mass: 72976.688 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Details: This a zinc-finger containing protein. Source: (natural) References: UniProt: P53165 |
-Non-polymers , 1 types, 8 molecules 
| #13: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn |
|---|
-Details
| Has ligand of interest | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.25 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 / Details: Solution were made from stock solution | ||||||||||||||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.2 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K / Details: blot for 4 seconds before plunging |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Cs: 2.7 mm / C2 aperture diameter: 70 µm / Alignment procedure: COMA FREE |
| Specimen holder | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 9 sec. / Electron dose: 44.17 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 313986 | ||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 113856 / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||
| Atomic model building | B value: 114.6 / Protocol: RIGID BODY FIT / Space: REAL / Target criteria: Correlation coefficient | ||||||||||||||||||||||||||||
| Atomic model building | PDB-ID: 4ZUX Accession code: 4ZUX / Source name: PDB / Type: experimental model |