 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6rtx | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
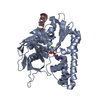
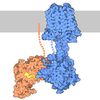
| Title | Crystal structure of the Patched-1 (PTCH1) ectodomain 1 | ||||||||||||
 Components Components | Protein patched homolog 1 | ||||||||||||
 Keywords Keywords | MEMBRANE PROTEIN / ---- Hedgehog morphogen receptor / receptor-nanobody complex / cholesterol / palmitate / lipid-protein-modification | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationneural plate axis specification / hedgehog receptor activity / neural tube patterning / smoothened binding / hedgehog family protein binding / Ligand-receptor interactions / neural tube formation / prostate gland development / limb morphogenesis / negative regulation of cell division ...neural plate axis specification / hedgehog receptor activity / neural tube patterning / smoothened binding / hedgehog family protein binding / Ligand-receptor interactions / neural tube formation / prostate gland development / limb morphogenesis / negative regulation of cell division / patched binding / somite development / Activation of SMO / smooth muscle tissue development / pharyngeal system development / cellular response to cholesterol / metanephric collecting duct development / response to alkaloid / commissural neuron axon guidance / regulation of smoothened signaling pathway / dorsal/ventral pattern formation / embryonic limb morphogenesis / Class B/2 (Secretin family receptors) / negative regulation of multicellular organism growth / cholesterol binding / ciliary membrane / dendritic growth cone / spermatid development / positive regulation of cholesterol efflux / response to retinoic acid / response to mechanical stimulus / negative regulation of osteoblast differentiation / axonal growth cone / Hedgehog 'off' state / liver regeneration / cyclin binding / animal organ morphogenesis / protein localization to plasma membrane / negative regulation of smoothened signaling pathway / brain development / protein processing / caveola / Hedgehog 'on' state / apical part of cell / endocytic vesicle membrane / response to estradiol / regulation of protein localization / heparin binding / midbody / postsynaptic membrane / response to xenobiotic stimulus / positive regulation of DNA-templated transcription / protein-containing complex binding / perinuclear region of cytoplasm / negative regulation of transcription by RNA polymerase II / signal transduction / plasma membrane Similarity search - Function | ||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.95 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.95 Å | ||||||||||||
 Authors Authors | Rudolf, A.F. / Kowatsch, C. / El Omari, K. / Malinauskas, T. / Kinnebrew, M. / Ansell, T.B. / Bishop, B. / Pardon, E. / Schwab, R.A. / Qian, M. ...Rudolf, A.F. / Kowatsch, C. / El Omari, K. / Malinauskas, T. / Kinnebrew, M. / Ansell, T.B. / Bishop, B. / Pardon, E. / Schwab, R.A. / Qian, M. / Duman, R. / Covey, D.F. / Steyaert, J. / Wagner, A. / Sansom, M.S.P. / Rohatgi, R. / Siebold, C. | ||||||||||||
| Funding support |  United Kingdom, 3items United Kingdom, 3items
| ||||||||||||
 Citation Citation | Journal: Nat Chem Biol / Year: 2019 Title: The morphogen Sonic hedgehog inhibits its receptor Patched by a pincer grasp mechanism. Authors: Amalie F Rudolf / Maia Kinnebrew / Christiane Kowatsch / T Bertie Ansell / Kamel El Omari / Benjamin Bishop / Els Pardon / Rebekka A Schwab / Tomas Malinauskas / Mingxing Qian / Ramona Duman ...Authors: Amalie F Rudolf / Maia Kinnebrew / Christiane Kowatsch / T Bertie Ansell / Kamel El Omari / Benjamin Bishop / Els Pardon / Rebekka A Schwab / Tomas Malinauskas / Mingxing Qian / Ramona Duman / Douglas F Covey / Jan Steyaert / Armin Wagner / Mark S P Sansom / Rajat Rohatgi / Christian Siebold /   Abstract: Hedgehog (HH) ligands, classical morphogens that pattern embryonic tissues in all animals, are covalently coupled to two lipids-a palmitoyl group at the N terminus and a cholesteroyl group at the C ...Hedgehog (HH) ligands, classical morphogens that pattern embryonic tissues in all animals, are covalently coupled to two lipids-a palmitoyl group at the N terminus and a cholesteroyl group at the C terminus. While the palmitoyl group binds and inactivates Patched 1 (PTCH1), the main receptor for HH ligands, the function of the cholesterol modification has remained mysterious. Using structural and biochemical studies, along with reassessment of previous cryo-electron microscopy structures, we find that the C-terminal cholesterol attached to Sonic hedgehog (Shh) binds the first extracellular domain of PTCH1 and promotes its inactivation, thus triggering HH signaling. Molecular dynamics simulations show that this interaction leads to the closure of a tunnel through PTCH1 that serves as the putative conduit for sterol transport. Thus, Shh inactivates PTCH1 by grasping its extracellular domain with two lipidic pincers, the N-terminal palmitate and the C-terminal cholesterol, which are both inserted into the PTCH1 protein core. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6rtx.cif.gz | 72.7 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6rtx.ent.gz | 51.7 KB | Display | PDB format |
| PDBx/mmJSON format | 6rtx.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/rt/6rtxftp://data.pdbj.org/pub/pdb/validation_reports/rt/6rtx | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  6rtwC  6rtySC  6rvcC  6rvdC S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj

- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 34670.301 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: Human PTCH1-ECD1 residues 139-428, containing three N-terminal additional residues derived from the expression vector pHLsec, and a C-terminal His6-Tag for IMAC purification Source: (gene. exp.) Homo sapiens (human) / Gene: PTCH1, PTCH / Plasmid: pHLsec / Cell line (production host): HEK293T / Production host: Homo sapiens (human) / References: UniProt: Q13635 | ||||
|---|---|---|---|---|---|
| #2: Sugar | 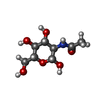  Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 2 Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 2Source method: isolated from a genetically manipulated source Formula: C8H15NO6 #3: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 99 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 99 / Source method: isolated from a natural source / Formula: H2OHas protein modification | Y | |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.37 Å3/Da / Density % sol: 48.14 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, sitting drop / pH: 8.5 Details: 0.1 M bicine/Trizma base pH 8.5, 0.03M of each ethylene glycol (diethyleneglycol, triethyleneglycol, tetraethyleneglycol and pentaethyleneglycol), 12.5% (v/v) MPD, 12.5% (w/v) PEG 1000, 12.5% (w/v) PEG 3350 |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Diamond / Beamline: I04 / Wavelength: 0.9795 Å |
| Detector | Type: DECTRIS PILATUS 6M / Detector: PIXEL / Date: Dec 15, 2016 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9795 Å / Relative weight: 1 |
| Reflection | Resolution: 1.95→50.14 Å / Num. obs: 24282 / % possible obs: 100 % / Redundancy: 6.4 % / CC1/2: 0.998 / Rmerge(I) obs: 0.129 / Rpim(I) all: 0.055 / Net I/σ(I): 10.7 |
| Reflection shell | Resolution: 1.95→1.99 Å / Redundancy: 5.7 % / Mean I/σ(I) obs: 1.4 / Num. unique obs: 1168 / CC1/2: 0.316 / Rpim(I) all: 0.525 / % possible all: 99.1 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 6RTY Resolution: 1.95→50.14 Å / Cor.coef. Fo:Fc: 0.958 / Cor.coef. Fo:Fc free: 0.93 / SU B: 5.484 / SU ML: 0.144 / Cross valid method: THROUGHOUT / ESU R: 0.158 / ESU R Free: 0.161 / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 36.903 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 1.95→50.14 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|