 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-6r23: The structure of a Ty3 retrotransposon capsid C-terminal domain dimer -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6r23 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | The structure of a Ty3 retrotransposon capsid C-terminal domain dimer | |||||||||
 Components Components | Transposon Ty3-I Gag-Pol polyprotein | |||||||||
 Keywords Keywords | VIRUS LIKE PARTICLE / Ty3 / retrotransposon / retrovirus / capsid / Gag polyprotein / capsid-CTD | |||||||||
| Function / homology |  Function and homology information Function and homology informationHydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases / ribonuclease H / DNA integration / RNA-directed DNA polymerase / RNA-directed DNA polymerase activity / RNA-DNA hybrid ribonuclease activity / DNA recombination / DNA-directed DNA polymerase / aspartic-type endopeptidase activity / DNA-directed DNA polymerase activity ...Hydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases / ribonuclease H / DNA integration / RNA-directed DNA polymerase / RNA-directed DNA polymerase activity / RNA-DNA hybrid ribonuclease activity / DNA recombination / DNA-directed DNA polymerase / aspartic-type endopeptidase activity / DNA-directed DNA polymerase activity / viral translational frameshifting / proteolysis / DNA binding / RNA binding / zinc ion binding / ATP binding / nucleus / cytoplasm Similarity search - Function | |||||||||
| Biological species |  | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.9 Å | |||||||||
 Authors Authors | Dodonova, S.O. / Prinz, S. / Bilanchone, V. / Sandmeyer, S. / Briggs, J.A.G. | |||||||||
| Funding support |  Germany, Germany,  United Kingdom, 2items United Kingdom, 2items
| |||||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2019 Title: Structure of the Ty3/Gypsy retrotransposon capsid and the evolution of retroviruses. Authors: Svetlana O Dodonova / Simone Prinz / Virginia Bilanchone / Suzanne Sandmeyer / John A G Briggs /  Abstract: Retroviruses evolved from long terminal repeat (LTR) retrotransposons by acquisition of envelope functions, and subsequently reinvaded host genomes. Together, endogenous retroviruses and LTR ...Retroviruses evolved from long terminal repeat (LTR) retrotransposons by acquisition of envelope functions, and subsequently reinvaded host genomes. Together, endogenous retroviruses and LTR retrotransposons represent major components of animal, plant, and fungal genomes. Sequences from these elements have been exapted to perform essential host functions, including placental development, synaptic communication, and transcriptional regulation. They encode a Gag polypeptide, the capsid domains of which can oligomerize to form a virus-like particle. The structures of retroviral capsids have been extensively described. They assemble an immature viral particle through oligomerization of full-length Gag. Proteolytic cleavage of Gag results in a mature, infectious particle. In contrast, the absence of structural data on LTR retrotransposon capsids hinders our understanding of their function and evolutionary relationships. Here, we report the capsid morphology and structure of the archetypal Gypsy retrotransposon Ty3. We performed electron tomography (ET) of immature and mature Ty3 particles within cells. We found that, in contrast to retroviruses, these do not change size or shape upon maturation. Cryo-ET and cryo-electron microscopy of purified, immature Ty3 particles revealed an irregular fullerene geometry previously described for mature retrovirus core particles and a tertiary and quaternary arrangement of the capsid (CA) C-terminal domain within the assembled capsid that is conserved with mature HIV-1. These findings provide a structural basis for studying retrotransposon capsids, including those domesticated in higher organisms. They suggest that assembly via a structurally distinct immature capsid is a later retroviral adaptation, while the structure of mature assembled capsids is conserved between LTR retrotransposons and retroviruses. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6r23.cif.gz | 52 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6r23.ent.gz | 33.5 KB | Display | PDB format |
| PDBx/mmJSON format | 6r23.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/r2/6r23ftp://data.pdbj.org/pub/pdb/validation_reports/r2/6r23 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4708MC  4707C  4709C  6r22C  6r24C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 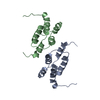
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 36382.371 Da / Num. of mol.: 2 / Mutation: D336I Source method: isolated from a genetically manipulated source Details: D336I mutation in the active center of the Ty3 protease Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: TY3B-I, YILWTy3-1 POL, YIL082W-A / Plasmid: pJK776 / Production host: Strain (production host): strain yVB1680 strain - BY4741 killer minus, Ty3 null References: UniProt: Q7LHG5, Hydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases, RNA-directed DNA polymerase, DNA-directed DNA polymerase, ribonuclease H |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Ty3 capsid C-terminal domain dimer / Type: COMPLEX Details: The 9 non-symmetry related copies of the Ty3 Capsid-CTD from the complete capsid shell structure were aligned and averaged to generate a higher resolution 4.9 A structure Entity ID: all / Source: RECOMBINANT | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight |
| ||||||||||||||||||||
| Source (natural) | Organism: | ||||||||||||||||||||
| Source (recombinant) | Organism: | ||||||||||||||||||||
| Details of virus | Empty: NO / Enveloped: NO / Isolate: OTHER / Type: VIRUS-LIKE PARTICLE | ||||||||||||||||||||
| Natural host | Organism: Saccharomyces cerevisiae | ||||||||||||||||||||
| Virus shell | Name: GAG Capsid / Diameter: 480 nm / Triangulation number (T number): 9 | ||||||||||||||||||||
| Buffer solution | pH: 8 | ||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid type: C-flat | ||||||||||||||||||||
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE / Humidity: 85 % / Chamber temperature: 296 K Details: The sample was applied onto glow-discharged C-flat (Protochips Inc.) holey carbon grids. The grids were blotted from the back side for 11 seconds at room temperature in a chamber at 85% ...Details: The sample was applied onto glow-discharged C-flat (Protochips Inc.) holey carbon grids. The grids were blotted from the back side for 11 seconds at room temperature in a chamber at 85% humidity and plunge-frozen into liquid ethane using a manual plunger. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS Details: Cryo-grids containing purified PR- Ty3 particles were imaged in a Titan Krios electron microscope equipped with a Falcon II direct electron detector, operated at 300 kV. Images were ...Details: Cryo-grids containing purified PR- Ty3 particles were imaged in a Titan Krios electron microscope equipped with a Falcon II direct electron detector, operated at 300 kV. Images were collected with a nominal magnification of 75000, giving a pixel size of 1.08 A. Images were collected in integrating mode with a total electron dose of 20 e/A2. The range of applied defocus values was between -1.0 um and -3.5 um. |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 75000 X / Nominal defocus max: 3500 nm / Nominal defocus min: 1000 nm / Cs: 2.7 mm / C2 aperture diameter: 50 µm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 20 e/Å2 / Detector mode: INTEGRATING / Film or detector model: FEI FALCON II (4k x 4k) / Num. of grids imaged: 1 |
| Image scans | Movie frames/image: 1 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 1727 | ||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.9 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 74160 Details: The Ty3 particles have a T=9 and each asymmetric unit of the lattice contains nine non-symmetry related Ty3 capsid protein monomers. We extracted all non-symmetry related units from the ...Details: The Ty3 particles have a T=9 and each asymmetric unit of the lattice contains nine non-symmetry related Ty3 capsid protein monomers. We extracted all non-symmetry related units from the final reconstructions from the two half datasets. The CA-CTDs were aligned and averaged separately. The final structure of the CA-CTD comes from nine averaged copies of the domain. The final structure of the CA-CTD was resolved at 4.9 A resolution at 0.143 FSC. Gaussian-smoothened ellipsoid-shaped masks were used for resolution measurements. Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||
| EM volume selection | Details: 1733 vesicles and near-complete buds were picked from 61 tomograms. Subtomograms were extracted from the surface of the vesicles. Num. of tomograms: 54 / Num. of volumes extracted: 2547 | ||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: Cross-correlation coefficient |
