Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
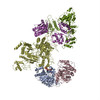
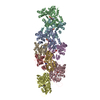
Yorodumi- PDB-6nzd: Cryo-EM Structure of the Lysosomal Folliculin Complex (FLCN-FNIP2... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6nzd | ||||||
|---|---|---|---|---|---|---|---|
| Title | Cryo-EM Structure of the Lysosomal Folliculin Complex (FLCN-FNIP2-RagA-RagC-Ragulator) | ||||||
 Components Components |
| ||||||
 Keywords Keywords | signaling protein/inhibitor / Lysosome / mTORC1 regulation / Amino acid sensing / GTPase / SIGNALING PROTEIN / signaling protein-inhibitor complex | ||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of cell proliferation involved in kidney development / cell proliferation involved in kidney development / negative regulation of post-translational protein modification / regulation of pro-B cell differentiation / regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / regulation of cholesterol efflux / Gtr1-Gtr2 GTPase complex / FNIP-folliculin RagC/D GAP ...negative regulation of cell proliferation involved in kidney development / cell proliferation involved in kidney development / negative regulation of post-translational protein modification / regulation of pro-B cell differentiation / regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / regulation of cholesterol efflux / Gtr1-Gtr2 GTPase complex / FNIP-folliculin RagC/D GAP / Ragulator complex / regulation of Ras protein signal transduction / regulation of TORC1 signaling / negative regulation of lysosome organization / protein localization to lysosome / lysosome localization / MTOR signalling / regulation of TOR signaling / ATPase inhibitor activity / Energy dependent regulation of mTOR by LKB1-AMPK / endosome organization / Amino acids regulate mTORC1 / negative regulation of brown fat cell differentiation / negative regulation of TOR signaling / cell-cell junction assembly / negative regulation of Rho protein signal transduction / kinase activator activity / negative regulation of cold-induced thermogenesis / negative regulation of glycolytic process / protein localization to membrane / lysosome organization / azurophil granule membrane / endosomal transport / regulation of cell size / small GTPase-mediated signal transduction / Macroautophagy / positive regulation of transforming growth factor beta receptor signaling pathway / RHOJ GTPase cycle / RHOQ GTPase cycle / TORC1 signaling / regulation of receptor recycling / CDC42 GTPase cycle / tertiary granule membrane / hemopoiesis / RHOG GTPase cycle / TOR signaling / response to amino acid / RHOH GTPase cycle / mTORC1-mediated signalling / ficolin-1-rich granule membrane / RAC3 GTPase cycle / RAC2 GTPase cycle / positive regulation of TOR signaling / enzyme-substrate adaptor activity / ERK1 and ERK2 cascade / energy homeostasis / specific granule membrane / epithelial cell proliferation / centriolar satellite / positive regulation of intrinsic apoptotic signaling pathway / RAC1 GTPase cycle / protein-membrane adaptor activity / positive regulation of TORC1 signaling / transforming growth factor beta receptor signaling pathway / positive regulation of autophagy / cellular response to nutrient levels / intrinsic apoptotic signaling pathway / guanyl-nucleotide exchange factor activity / cholesterol homeostasis / RNA splicing / viral genome replication / enzyme inhibitor activity / cellular response to amino acid starvation / negative regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / cellular response to starvation / Regulation of PTEN gene transcription / tumor necrosis factor-mediated signaling pathway / negative regulation of autophagy / positive regulation of interleukin-8 production / GTPase activator activity / TP53 Regulates Metabolic Genes / cellular response to amino acid stimulus / phosphatidylinositol 3-kinase/protein kinase B signal transduction / phosphoprotein binding / regulation of cell growth / positive regulation of protein-containing complex assembly / negative regulation of ERK1 and ERK2 cascade / MAP2K and MAPK activation / positive regulation of protein localization to nucleus / negative regulation of epithelial cell proliferation / intrinsic apoptotic signaling pathway in response to DNA damage / response to virus / mitotic spindle / intracellular protein localization / GDP binding / late endosome / glucose homeostasis / late endosome membrane / E3 ubiquitin ligases ubiquitinate target proteins / in utero embryonic development Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.6 Å | ||||||
 Authors Authors | Fromm, S.A. / Young, L.N. / Hurley, J.H. | ||||||
| Funding support |  United States, 1items United States, 1items
| ||||||
 Citation Citation | Journal: Science / Year: 2019 Title: Structural mechanism of a Rag GTPase activation checkpoint by the lysosomal folliculin complex. Authors: Rosalie E Lawrence / Simon A Fromm / Yangxue Fu / Adam L Yokom / Do Jin Kim / Ashley M Thelen / Lindsey N Young / Chun-Yan Lim / Avi J Samelson / James H Hurley / Roberto Zoncu / Abstract: The tumor suppressor folliculin (FLCN) enables nutrient-dependent activation of the mechanistic target of rapamycin complex 1 (mTORC1) protein kinase via its guanosine triphosphatase (GTPase) ...The tumor suppressor folliculin (FLCN) enables nutrient-dependent activation of the mechanistic target of rapamycin complex 1 (mTORC1) protein kinase via its guanosine triphosphatase (GTPase) activating protein (GAP) activity toward the GTPase RagC. Concomitant with mTORC1 inactivation by starvation, FLCN relocalizes from the cytosol to lysosomes. To determine the lysosomal function of FLCN, we reconstituted the human lysosomal FLCN complex (LFC) containing FLCN, its partner FLCN-interacting protein 2 (FNIP2), and the RagA:RagC GTPases as they exist in the starved state with their lysosomal anchor Ragulator complex and determined its cryo-electron microscopy structure to 3.6 angstroms. The RagC-GAP activity of FLCN was inhibited within the LFC, owing to displacement of a catalytically required arginine in FLCN from the RagC nucleotide. Disassembly of the LFC and release of the RagC-GAP activity of FLCN enabled mTORC1-dependent regulation of the master regulator of lysosomal biogenesis, transcription factor E3, implicating the LFC as a checkpoint in mTORC1 signaling. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6nzd.cif.gz | 357.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6nzd.ent.gz | 259.6 KB | Display | PDB format |
| PDBx/mmJSON format | 6nzd.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/nz/6nzdftp://data.pdbj.org/pub/pdb/validation_reports/nz/6nzd | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  0554MC  0556C C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj





- Assembly
Assembly
| Deposited unit | 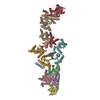
|
|---|---|
| 1 |
|
-Components
-Ragulator complex protein ... , 4 types, 4 molecules ABCD
| #1: Protein | Mass: 18325.350 Da / Num. of mol.: 1 / Mutation: G2A Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LAMTOR1, C11orf59, PDRO, PP7157 / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: Q6IAA8 Spodoptera frugiperda (fall armyworm) / References: UniProt: Q6IAA8 |
|---|---|
| #2: Protein | Mass: 13645.579 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LAMTOR2, MAPBPIP, ROBLD3, HSPC003 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9Y2Q5 |
| #3: Protein | Mass: 13637.678 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LAMTOR3, MAP2K1IP1, MAPKSP1, PRO2783 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9UHA4 |
| #4: Protein | Mass: 10753.236 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LAMTOR4, C7orf59 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q0VGL1 |
-Protein , 3 types, 3 molecules EHI
| #5: Protein | Mass: 18178.520 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LAMTOR5, HBXIP, hCG_40252 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: A0A0C4DGV4, UniProt: O43504*PLUS |
|---|---|
| #8: Protein | Mass: 69143.070 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: FLCN, BHD / Cell line (production host): HEK 293 GNTI / Production host: Homo sapiens (human) / References: UniProt: Q8NFG4 |
| #9: Protein | Mass: 122475.414 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: FNIP2, FNIPL, KIAA1450, MAPO1 / Cell line (production host): HEK 293 GNTI / Production host: Homo sapiens (human) / References: UniProt: Q9P278 |
-Ras-related GTP-binding protein ... , 2 types, 2 molecules FG
| #6: Protein | Mass: 36615.168 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RRAGA / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q7L523 |
|---|---|
| #7: Protein | Mass: 44758.336 Da / Num. of mol.: 1 / Mutation: D181N Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RRAGC / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9HB90 |
-Non-polymers , 2 types, 2 molecules 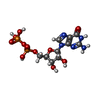

| #10: Chemical | ChemComp-GDP /  Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM |
|---|---|
| #11: Chemical | ChemComp-L8S /  Mass: 540.231 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N4O14P3S Mass: 540.231 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N4O14P3S |
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: FLCN-FNIP2-RagA-RagC-Ragulator Complex / Type: COMPLEX / Details: RagA bound to GDP; RagC bound to XTPgammaS / Entity ID: #1-#9 / Source: MULTIPLE SOURCES | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.34 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||
| Source (natural) | Organism: Homo sapiens (human) | ||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: Spodoptera frugiperda (fall armyworm) | ||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.4 | ||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.5 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Specimen support | Details: 15 W / Grid material: COPPER / Grid mesh size: 300 divisions/in. / Grid type: C-flat-2/1 | ||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 90 % / Chamber temperature: 277 K / Details: Whatman 597 |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 3000 nm / Nominal defocus min: 1000 nm / Cs: 2.7 mm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 11 sec. / Electron dose: 65.6 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 1 / Num. of real images: 2703 |
| EM imaging optics | Energyfilter slit width: 20 eV |
| Image scans | Movie frames/image: 44 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 982343 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 163376 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Space: REAL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | 3D fitting-ID: 1 / Source name: PDB / Type: experimental model
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Highest resolution: 3.6 Å |