 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-6h82: Cryo-EM structure of the archaeal extremophilic internal membrane... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6h82 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of the archaeal extremophilic internal membrane containing Haloarcula hispanica icosahedral virus 2 (HHIV-2) at 3.78 Angstroms resolution. | |||||||||||||||
 Components Components |
| |||||||||||||||
 Keywords Keywords | VIRUS / single vertical beta-barrel virus / archaeal / membrane-containing / quasi-atomic resolution | |||||||||||||||
| Function / homology | VP7 / VP4 / Uncharacterized protein Function and homology information Function and homology information | |||||||||||||||
| Biological species |  Haloarcula hispanica icosahedral virus 2 Haloarcula hispanica icosahedral virus 2 | |||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.78 Å | |||||||||||||||
 Authors Authors | Abrescia, N.G. / Santos-Perez, I. / Charro, D. | |||||||||||||||
| Funding support |  Spain, Spain,  Finland, 4items Finland, 4items
| |||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2019 Title: Structural basis for assembly of vertical single β-barrel viruses. Authors: Isaac Santos-Pérez / Diego Charro / David Gil-Carton / Mikel Azkargorta / Felix Elortza / Dennis H Bamford / Hanna M Oksanen / Nicola G A Abrescia / Abstract: The vertical double β-barrel major capsid protein (MCP) fold, fingerprint of the PRD1-adeno viral lineage, is widespread in many viruses infecting organisms across the three domains of life. The ...The vertical double β-barrel major capsid protein (MCP) fold, fingerprint of the PRD1-adeno viral lineage, is widespread in many viruses infecting organisms across the three domains of life. The discovery of PRD1-like viruses with two MCPs challenged the known assembly principles. Here, we present the cryo-electron microscopy (cryo-EM) structures of the archaeal, halophilic, internal membrane-containing Haloarcula californiae icosahedral virus 1 (HCIV-1) and Haloarcula hispanica icosahedral virus 2 (HHIV-2) at 3.7 and 3.8 Å resolution, respectively. Our structures reveal proteins located beneath the morphologically distinct two- and three-tower capsomers and homopentameric membrane proteins at the vertices that orchestrate the positioning of pre-formed vertical single β-barrel MCP heterodimers. The cryo-EM based structures together with the proteomics data provide insights into the assembly mechanism of this type of viruses and into those with membrane-less double β-barrel MCPs. #1: Journal: Structure / Year: 2015Title: Insight into the Assembly of Viruses with Vertical Single β-barrel Major Capsid Proteins. Authors: David Gil-Carton / Salla T Jaakkola / Diego Charro / Bibiana Peralta / Daniel Castaño-Díez / Hanna M Oksanen / Dennis H Bamford / Nicola G A Abrescia /  Abstract: Archaeal viruses constitute the least explored niche within the virosphere. Structure-based approaches have revealed close relationships between viruses infecting organisms from different domains of ...Archaeal viruses constitute the least explored niche within the virosphere. Structure-based approaches have revealed close relationships between viruses infecting organisms from different domains of life. Here, using biochemical and cryo-electron microscopy techniques, we solved the structure of euryarchaeal, halophilic, internal membrane-containing Haloarcula hispanica icosahedral virus 2 (HHIV-2). We show that the density of the two major capsid proteins (MCPs) recapitulates vertical single β-barrel proteins and that disulfide bridges stabilize the capsid. Below, ordered density is visible close to the membrane and at the five-fold vertices underneath the host-interacting vertex complex underpinning membrane-protein interactions. The HHIV-2 structure exemplifies the division of conserved architectural elements of a virion, such as the capsid, from those that evolve rapidly due to selective environmental pressure such as host-recognizing structures. We propose that in viruses with two vertical single β-barrel MCPs the vesicle is indispensable, and membrane-protein interactions serve as protein-railings for guiding the assembly. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
 UCSF Chimera
UCSF Chimera- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6h82.cif.gz | 1.7 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6h82.ent.gz | 1.5 MB | Display | PDB format |
| PDBx/mmJSON format | 6h82.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/h8/6h82ftp://data.pdbj.org/pub/pdb/validation_reports/h8/6h82 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  0172MC  0050C  0072C  0073C 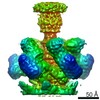 0131C  0174C  6h9cC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj- Assembly
Assembly
| Deposited unit | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | x 60
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS oper:
|