 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5vey | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
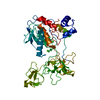
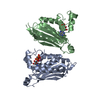
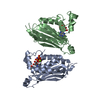
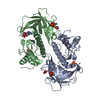
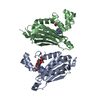
| タイトル | Solution NMR structure of histone H2A-H2B mono-ubiquitylated at H2A Lys15 in complex with RNF169 (653-708) | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | TRANSFERASE / STRUCTURAL PROTEIN / SIGNALING PROTEIN / Nucleosome / Ubiquitin / Ubiquitin ligase / Complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報negative regulation of double-strand break repair / ubiquitin-modified histone reader activity / hypothalamus gonadotrophin-releasing hormone neuron development / female meiosis I / positive regulation of protein monoubiquitination / fat pad development / mitochondrion transport along microtubule / K63-linked polyubiquitin modification-dependent protein binding / seminiferous tubule development / female gonad development ...negative regulation of double-strand break repair / ubiquitin-modified histone reader activity / hypothalamus gonadotrophin-releasing hormone neuron development / female meiosis I / positive regulation of protein monoubiquitination / fat pad development / mitochondrion transport along microtubule / K63-linked polyubiquitin modification-dependent protein binding / seminiferous tubule development / female gonad development / male meiosis I / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / negative regulation of tumor necrosis factor-mediated signaling pathway / nucleosome binding / neuron projection morphogenesis / protein localization to CENP-A containing chromatin / energy homeostasis / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / regulation of proteasomal protein catabolic process / Packaging Of Telomere Ends / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / IRAK2 mediated activation of TAK1 complex / Prevention of phagosomal-lysosomal fusion / Alpha-protein kinase 1 signaling pathway / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Negative regulation of FLT3 / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Deposition of new CENPA-containing nucleosomes at the centromere / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Regulation of FZD by ubiquitination / Downregulation of ERBB4 signaling / APC-Cdc20 mediated degradation of Nek2A / p75NTR recruits signalling complexes / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / regulation of neuron apoptotic process / NF-kB is activated and signals survival / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / RNA Polymerase I Promoter Opening / Pexophagy / Regulation of innate immune responses to cytosolic DNA / NRIF signals cell death from the nucleus / Downregulation of ERBB2:ERBB3 signaling / Regulation of PTEN localization / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / VLDLR internalisation and degradation / Meiotic synapsis / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / positive regulation of protein ubiquitination / Translesion synthesis by REV1 / Regulation of BACH1 activity / TICAM1, RIP1-mediated IKK complex recruitment / Translesion synthesis by POLK / InlB-mediated entry of Listeria monocytogenes into host cell / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / DNA methylation / Downregulation of TGF-beta receptor signaling / Condensation of Prophase Chromosomes / Josephin domain DUBs / Translesion synthesis by POLI / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / IKK complex recruitment mediated by RIP1 / Gap-filling DNA repair synthesis and ligation in GG-NER / PINK1-PRKN Mediated Mitophagy / regulation of mitochondrial membrane potential / Regulation of activated PAK-2p34 by proteasome mediated degradation / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / TNFR1-induced NF-kappa-B signaling pathway / PRC2 methylates histones and DNA / innate immune response in mucosa / TCF dependent signaling in response to WNT 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 溶液NMR / simulated annealing | |||||||||
 データ登録者 データ登録者 | Hu, Q. / Botuyan, M.V. / Cui, G. / Mer, G. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Mol. Cell / 年: 2017 タイトル: Mechanisms of Ubiquitin-Nucleosome Recognition and Regulation of 53BP1 Chromatin Recruitment by RNF168/169 and RAD18. 著者: Hu, Q. / Botuyan, M.V. / Cui, G. / Zhao, D. / Mer, G. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5vey.cif.gz | 1.9 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5vey.ent.gz | 1.6 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5vey.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ve/5veyftp://data.pdbj.org/pub/pdb/validation_reports/ve/5vey | HTTPS FTP |
|---|
-関連構造データ
| 関連構造データ |  5vf0C C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ | |
| その他のデータベース |
|





-リンク
 PDBj
PDBj
























- 集合体
集合体
| 登録構造単位 | 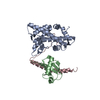
| |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| |||||||||
| NMR アンサンブル |
|
 gel filtration
gel filtration-要素
| #1: タンパク質 | 分子量: 21658.943 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H2BJ, H2BFR, HIST1H2AB, H2AFM, HIST1H2AE, H2AFA プラスミド: pT7.7 / 発現宿主:  |
|---|---|
| #2: タンパク質 | 分子量: 8576.831 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: UBB / プラスミド: pET / 発現宿主: |
| #3: タンパク質 | 分子量: 7016.938 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RNF169, KIAA1991 / プラスミド: pTEV / 発現宿主: |
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 溶液NMR | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NMR実験 |
|